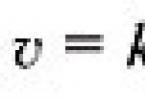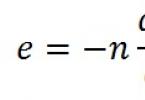Lipidy resyntetizované v čreve sú transportované ako súčasť chylomikrónov s lymfou, lipidy sú nerozpustné vo vode, preto sú transportované v spojení s bielkovinami.
Lipoproteíny sú komplexy proteínov a lipidov, transportná forma lipidov v krvi. LP lipidy: triglyceridy, fosfolipidy, cholesterol. LP proteíny sú apoproteíny jednotlivých tried LP.
Funkcie apoproteínov sú štrukturálne (LP), transportné, sekrečné (potrebné pre sekréciu LP pečeňovými a črevnými bunkami), nevyhnutné pre interakciu LP s receptormi, aktivujú enzýmy podieľajúce sa na metabolizme LP, prepožičiavajú lipidom rozpustnosť vo vode. apoproteín A1 v HDL aktivuje LCAT.
Lipoproteínová štruktúra. Hydrofóbne jadro (estery cholesterolu, TG) je zvonka obklopené fosfolipidmi, voľným cholesterolom a apoproteínmi.
Klasifikácia liekov.
na základe mobility v elektrickom poli: - ChM zostávajú na štarte, - ostatné migrujú do globulínových zón: ß-LP, pre-ß-LP, α-LP.
Podľa hydratovanej hustoty (metódou ultracentrifugácie) sa liečivá delia na CM, VLDL, LDPP, LDL a HDL.
Biologická úloha liekov. Endogénne TG sa dodávajú do periférnych buniek na uspokojenie energetických potrieb a endogénny cholesterol sa dodáva do membránovej biosyntézy.
Zloženie a vlastnosti lipoproteínov.
Chylomikróny transportujú exogénne TG, cholesterol, fosfolipidy a tuky z potravy z čreva do tkanív cez lymfatický systém. Nezrelé KM sa syntetizujú v enterocytoch, ktoré sa najskôr dostanú do lymfy a potom do krvného obehu. Hlavný apoproteín KM, proteín B-48, je syntetizovaný v bunkách črevnej sliznice a je nevyhnutný pre tvorbu štruktúry KM. V krvi nezrelé KM prijímajú ďalšie apoproteíny z HDL - C- a E- a premieňajú sa na zrelé KM. Prvým orgánom, cez ktorý musí chemoterapia prejsť, sú pľúca. Keď KM vstúpi z čreva do krvného obehu, žírne bunky sa aktivujú uvoľnením heparínu a aktiváciou lipoproteínovej lipázy.
Adsorpčná lipémia je zvýšenie množstva lipidov v krvi, ku ktorému dochádza po jedle.
Lipoproteínová lipáza (clearing factor) hydrolyzuje TAG na CM a VLDL, nachádza sa v endoteli kapilár rôznych orgánov, je aktivovaná heparínom a zvýšením TAG v krvi. TAG chylomikróny sa štiepia na povrchu a vo vnútri hepatocytov, na povrchu endotelu kapilár tukového tkaniva.
VLDL a HDL sú vylučované do krvi pečeňou, kde sú syntetizované. LDL sa tvorí v krvnom obehu z VLDL ako výsledok hydrolýzy TG časti VLDL lipoproteínovou lipázou.
Osud LDL. Na plazmatických membránach buniek sú receptory pre LDL. LDL preniká do buniek, kde sa vplyvom lyzozomálnych hydroláz rozkladajú na svoje zložky, voľný cholesterol je obsiahnutý v plazmatickej membráne alebo sa esterifikuje a ukladá vo forme esterov v cytoplazme. Je možná nešpecifická endocytóza LDL.
HDL prenáša cholesterol do pečene. V pečeni sa cholesterol oxiduje na žlčové kyseliny a odstraňuje sa cez črevá. K oxidácii cholesterolu dochádza v pečeni systémom monooxygenázy. Cholesterol 7a-hydroxyláza je enzým obmedzujúci rýchlosť. HDL je schopný prijímať cholesterol z bunkových membrán.
Premena voľného cholesterolu na esterifikovaný: cholesterol + lecitín → lyzolecitín + ester cholesterolu. Ester cholesterolu sa tvorí na povrchu HDL a prenáša sa do jadra HDL.
Pokles HDL cholesterolu v krvnej plazme je spojený s poklesom LCAT, počtu HDL častíc, lecitínu a apoproteínu A1.
Polovičný život. HM – menej ako hodinu, VLDL – 2-4 hodiny, LDL – 2-4 dni, HDL – 5 dní. LDL a HDL sú absorbované endocytózou bunkami pečene, čriev, tukového tkaniva, obličiek a nadobličiek a sú zničené v lyzozómoch.
Neesterifikované mastné kyseliny (NEFA). Mastné kyseliny v krvnej plazme sú v esterifikovanej forme: v zložení fosfolipidov, esterov cholesterolu, mono-, di-, triglyceridov. Vo voľnej forme sú mastné kyseliny transportované v plazme z tukového tkaniva a pečene do kostrového svalstva, v tomto prípade sú viazané na albumín.
NEFA sa dostávajú do krvnej plazmy v dôsledku lipolýzy TG, katalyzovanej lipázou v tukovom tkanive, vznikajú pôsobením lipoproteínovej lipázy na TG krvnej plazmy pri ich prechode do tkanív, mastné kyseliny s dĺžkou reťazca menšou ako 10 atómov uhlíka sa absorbuje v neesterifikovanej forme cez portálny obehový systém a dostane sa do pečene (to je dôležité pre deti, pretože mlieko je bohaté na mastné kyseliny s krátkym reťazcom).
Triacylglyceridy sú transportnou formou pre nasýtené mastné kyseliny. Fosfolipidy a cholesterol sú transportnou formou pre polynenasýtené mastné kyseliny.
Funkcie NEFA - poskytujú 50% energie počas pôstu, energetický materiál pre myokard, svaly, obličky, pečeň, nasýtené mastné kyseliny plnia energiu a nenasýtené mastné kyseliny plnia plastické funkcie.
Odpoveď. Amoniak, glutamín a amónny ión NH 4 + sa zvyšujú v krvi pacienta.
Transport lipidov v tele prebieha dvoma spôsobmi:
- 1) mastné kyseliny sú transportované v krvi pomocou albumínov;
- 2) TG, FL, HS, EHS atď. Lipidy sú transportované krvou ako súčasť lipoproteínov.
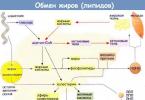
Metabolizmus lipoproteínov
Lipoproteíny (LP) sú sférické supramolekulárne komplexy pozostávajúce z lipidov, proteínov a sacharidov. LP majú hydrofilný obal a hydrofóbne jadro. Súčasťou hydrofilného obalu sú bielkoviny a amfifilné lipidy – PL, cholesterol. Hydrofóbne jadro zahŕňa hydrofóbne lipidy – TG, estery cholesterolu atď. LP sú vysoko rozpustné vo vode.
V tele sa syntetizuje niekoľko druhov lipidov, líšia sa chemickým zložením, tvoria sa na rôznych miestach a transportujú lipidy rôznymi smermi.
Lieky sa delia pomocou:
- 1) elektroforéza, podľa náboja a veľkosti, na b-LP, v-LP, pre-c-LP a CM;
- 2) centrifugácia podľa hustoty pre HDL, LDL, LDLP, VLDL a CM.
Pomer a množstvo LP v krvi závisí od dennej doby a výživy. Počas postabsorpčného obdobia a počas hladovania sú v krvi prítomné len LDL a HDL.
Hlavné typy lipoproteínov
Zloženie, % VLDL CM
- (pre-in-LP) DILI
- (pre-in-LP) LDL
- (v-LP) HDL
- (b-LP)
Bielkoviny 2 10 11 22 50
FL 3 18 23 21 27
EHS 3 10 30 42 16
TG 85 55 26 7 3
Hustota, g/ml 0,92-0,98 0,96-1,00 0,96-1,00 1,00-1,06 1,06-1,21
Priemer, nm >120 30-100 30-100 21-100 7-15
Funkcie Transport exogénnych potravinových lipidov do tkanív Transport endogénnych pečeňových lipidov do tkanív Transport endogénnych pečeňových lipidov do tkanív Transport cholesterolu
v tkanive Odstránenie prebytočného cholesterolu
z látok
apo A, C, E
Miesto vzniku enterocytového hepatocytu v krvi z VLDL v krvi z LDLP hepatocytu
Apo B-48, C-II, E B-100, C-II, E B-100, E B-100 A-I C-II, E, D
Normálne v krvi< 2,2 ммоль/л 0,9- 1,9 ммоль/л
Apobelki
Proteíny, ktoré tvoria liečivo, sa nazývajú apoproteíny (apoproteíny, apo). Medzi najčastejšie apoproteíny patria: apo A-I, A-II, B-48, B-100, C-I, C-II, C-III, D, E. Apoproteíny môžu byť periférne (hydrofilné: A-II, C-II, E) a integrálne (majú hydrofóbnu oblasť: B-48, B-100). Periférny apos sa prenáša medzi LP, ale integrálny apos nie. Apoproteíny vykonávajú niekoľko funkcií:
Funkcia apoproteínu Miesto vzniku Lokalizácia
A-I aktivátor LCAT, tvorba ECS pečeňového HDL
A-II Aktivátor LCAT, tvorba ECS HDL, CM
B-48 Štrukturálny (LP syntéza), receptorový (LP fagocytóza) enterocyt XM
B-100 Štrukturálne (LP syntéza), receptor (LP fagocytóza) pečeň VLDL, LDPP, LDL
C-I aktivátor LCAT, tvorba ECS Pečeň HDL, VLDL
C-II LPL aktivátor, stimuluje hydrolýzu TG v lipoproteíne Pečeň HDL > CM, VLDL
C-III inhibítor LPL, inhibuje hydrolýzu TG v LP Pečeň HDL > CM, VLDL
D Prenos esteru cholesterolu (CET) Pečeň HDL
E receptor, fagocytóza LP pečeň HDL > CM, VLDL, LDLP
Enzýmy transportujúce lipidy
Lipoproteínová lipáza (LPL) (EC 3.1.1.34, gén LPL, asi 40 defektných alel) je spojená s heparansulfátom lokalizovaným na povrchu endotelových buniek kapilár krvných ciev. Hydrolyzuje TG v zložení liečiva na glycerol a 3 mastné kyseliny. So stratou TG sa CM mení na zvyškový CM a VLDL zvyšuje svoju hustotu na LDLP a LDL.
Apo C-II LP aktivuje LPL a fosfolipidy LP sa podieľajú na väzbe LPL na povrch LP. Syntéza LPL je indukovaná inzulínom. Apo C-III inhibuje LPL.
LPL sa syntetizuje v bunkách mnohých tkanív: tuk, svaly, pľúca, slezina, bunky laktujúcej mliečnej žľazy. Nie je v pečeni. Izoenzýmy LPL rôznych tkanív sa líšia v hodnotách Km. V tukovom tkanive má LPL Km 10x väčšie ako v myokarde, preto tukové tkanivo absorbuje mastné kyseliny len pri nadbytku TG v krvi a myokard neustále aj pri nízkej koncentrácii TG v krvi. Mastné kyseliny v adipocytoch sa využívajú na syntézu TG, v myokarde ako zdroj energie.
Hepatálna lipáza sa nachádza na povrchu hepatocytov, nepôsobí na zrelý cholesterol, ale hydrolyzuje TG v LDPP.
Lecitín: cholesterolacyltransferáza (LCAT) sa nachádza v HDL, prenáša acyl z lecitínu na cholesterol za vzniku ECL a lyzolecitínu. Je aktivovaný apo A-I, A-II a C-I.
lecitín + CS > lyzolecitín + ECS
ECS sa ponorí do jadra HDL alebo sa prenesie za účasti apo D do iného HDL.
Receptory transportu lipidov
LDL receptor je komplexný proteín pozostávajúci z 5 domén a obsahujúci sacharidovú časť. LDL receptor interaguje s proteínmi ano B-100 a apo E, viaže LDL dobre, horšie ako DILI, VLDL a zvyškový CM obsahujúci tieto apos. Tkanivové bunky obsahujú na svojom povrchu veľké množstvo LDL receptorov. Napríklad jedna fibroblastová bunka má od 20 000 do 50 000 receptorov.
Ak množstvo cholesterolu vstupujúceho do bunky prevyšuje jeho potrebu, potom je potlačená syntéza LDL receptorov, čo znižuje tok cholesterolu z krvi do buniek. Pri znížení koncentrácie voľného cholesterolu v bunke sa naopak aktivuje syntéza HMG-CoA reduktázy a LDL receptorov. Hormóny stimulujú syntézu LDL receptorov: inzulín a trijódtyronín (T3), pohlavné hormóny a glukokortikoidy ju znižujú.
Proteín podobný LDL receptoru Na povrchu buniek v mnohých orgánoch (pečeň, mozog, placenta) existuje ďalší typ receptora nazývaný „proteín podobný LDL receptoru“. Tento receptor interaguje s apo E a zachytáva zvyšok (reziduálne) CM a DILI. Keďže zvyškové častice obsahujú cholesterol, tento typ receptorov zabezpečuje aj jeho vstup do tkanív.
Okrem vstupu cholesterolu do tkanív endocytózou lipoproteínov sa určité množstvo cholesterolu dostáva do buniek difúziou z LDL a iných lipoproteínov pri ich kontakte s bunkovými membránami.
Normálna koncentrácia v krvi je:
- * LDL
- * celkové lipidy 4-8g/l,
- * TG 0,5-2,1 mmol/l,
- * Voľné mastné kyseliny 400-800 µmol/l
Lipidy sú nerozpustné vo vodnom prostredí, preto pre ich transport v organizme vznikajú komplexy lipidov s proteínmi – lipoproteíny (LP). Existuje exo- a endogénny transport lipidov. Exogénna zahŕňa transport lipidov prijatých z potravy a endogénna zahŕňa pohyb lipidov syntetizovaných v tele.
Existuje niekoľko druhov LP, ale všetky majú podobnú štruktúru – hydrofóbne jadro a hydrofilnú vrstvu na povrchu. Hydrofilnú vrstvu tvoria proteíny nazývané apoproteíny a amfifilné molekuly lipidov – fosfolipidy a cholesterol. Hydrofilné skupiny týchto molekúl smerujú k vodnej fáze a hydrofóbne skupiny smerujú k jadru, v ktorom sa nachádzajú transportované lipidy. Apoproteíny vykonávajú niekoľko funkcií:
· tvoria štruktúru lipoproteínov (napr. B-48 je hlavný proteín XM, B-100 je hlavný proteín VLDL, LDPP, LDL);
· interagovať s receptormi na povrchu buniek, určiť, ktoré tkanivá zachytia tento typ lipoproteínu (apoproteín B-100, E);
· sú enzýmy alebo aktivátory enzýmov pôsobiacich na lipoproteíny (C-II - aktivátor lipoproteínovej lipázy, A-I - aktivátor lecitínu: cholesterolacyltransferáza).
Pri exogénnom transporte sa TAG resyntetizujú v enterocytoch spolu s fosfolipidmi, cholesterolom a proteínmi tvoria CM a v tejto forme sa vylučujú najskôr do lymfy a potom do krvi. V lymfe a krvi sa apoproteíny E (apo E) a C-II (apo C-II) prenášajú z HDL do CM, čím sa CM mení na „zrelé“. ChM sú dosť veľké, takže po zjedení mastného jedla dodajú krvnej plazme opalizujúci, mliečny vzhľad. Keď sa CM dostanú do obehového systému, rýchlo podstúpia katabolizmus a zmiznú v priebehu niekoľkých hodín. Čas deštrukcie KM závisí od hydrolýzy TAG pôsobením lipoproteínovej lipázy (LPL). Tento enzým je syntetizovaný a vylučovaný tukovým a svalovým tkanivom a bunkami mliečnych žliaz. Vylučovaný LPL sa viaže na povrch endotelových buniek kapilár tkanív, kde bol syntetizovaný. Regulácia sekrécie je špecifická pre tkanivo. V tukovom tkanive je syntéza LPL stimulovaná inzulínom. Tým je zabezpečený prísun mastných kyselín pre syntézu a skladovanie vo forme TAG. Pri diabetes mellitus, keď je nedostatok inzulínu, hladina LPL klesá. V dôsledku toho sa v krvi hromadí veľké množstvo LP. Vo svaloch, kde sa LPL podieľa na dodávaní mastných kyselín na oxidáciu medzi jedlami, inzulín inhibuje tvorbu tohto enzýmu.
Na povrchu CM sú 2 faktory nevyhnutné pre aktivitu LPL: apoC-II a fosfolipidy. ApoC-II aktivuje tento enzým a fosfolipidy sa podieľajú na väzbe enzýmu na povrch KM. V dôsledku pôsobenia LPL na molekuly TAG vznikajú mastné kyseliny a glycerol. Väčšina mastných kyselín preniká do tkanív, kde sa môžu ukladať vo forme TAG (tukové tkanivo) alebo využiť ako zdroj energie (svaly). Glycerol je krvou transportovaný do pečene, kde môže byť počas doby vstrebávania využitý na syntézu tukov.
V dôsledku pôsobenia LPL sa množstvo neutrálnych tukov v CM zníži o 90 %, veľkosť častíc sa zníži a apoC-II sa prenesie späť do HDL. Výsledné častice sa nazývajú zvyškové CM (zvyšky). Obsahujú PL, cholesterol, vitamíny rozpustné v tukoch, apoB-48 a apoE. Zvyškové KM sú zachytené hepatocytmi, ktoré majú receptory, ktoré interagujú s týmito apoproteínmi. Pôsobením lyzozómových enzýmov sú proteíny a lipidy hydrolyzované a následne využité. Vitamíny rozpustné v tukoch a exogénny cholesterol sa využívajú v pečeni alebo transportujú do iných orgánov.
Počas endogénneho transportu sú TAG a PL resyntetizované v pečeni zahrnuté do VLDL, ktorý zahŕňa apoB100 a apoC. VLDL je hlavnou transportnou formou pre endogénny TAG. Keď je VLDL v krvi, dostane apoC-II a apoE z HDL a je vystavený LPL. Počas tohto procesu sa VLDL najskôr premení na LDLP a potom na LDL. Hlavným lipidom LDL sa stáva cholesterol, ktorý sa v ich zložení prenáša do buniek všetkých tkanív. Mastné kyseliny vznikajúce pri hydrolýze vstupujú do tkanív a glycerol je krvou transportovaný do pečene, kde sa môže opäť použiť na syntézu TAG.
Všetky zmeny v obsahu liečiv v krvnej plazme, charakterizované ich zvýšením, znížením alebo úplnou absenciou, sa spájajú pod názvom dislipoproteinémia. Dyslipoproteinémia môže byť buď špecifickým primárnym prejavom porúch metabolizmu lipidov a lipoproteínov, alebo sprievodným syndrómom pri niektorých ochoreniach vnútorných orgánov (sekundárna dyslipoproteinémia). Pri úspešnej liečbe základného ochorenia zmiznú.
Hypolipoproteinémia zahŕňa nasledujúce stavy.
1. Abetalipoproteinémia vzniká v dôsledku zriedkavého dedičného ochorenia – defektu génu apoproteínu B, kedy je narušená syntéza proteínov apoB-100 v pečeni a apoB-48 v čreve. V dôsledku toho sa v bunkách črevnej sliznice netvoria CM a v pečeni sa netvorí VLDL a v bunkách týchto orgánov sa hromadia kvapky tuku.
2. Familiárna hypobetalipoproteinémia: koncentrácia liečiv s obsahom apoB je len 10-15% normálnej hladiny, ale telo je schopné tvoriť cholesterol.
3. Familiárny deficit a-LP (Tangierova choroba): v krvnej plazme sa prakticky nenachádza HDL a v tkanivách sa hromadí veľké množstvo esterov cholesterolu, pacientom chýba apoC-II, ktorý je aktivátorom LPL, čo vedie k zvýšenie koncentrácie TAG charakteristické pre tento stav v krvnej plazme.
Medzi hyperlipoproteinémiami sa rozlišujú nasledujúce typy.
Typ I - hyperchylomikroémia. Rýchlosť odstraňovania KM z krvného obehu závisí od aktivity LPL, prítomnosti HDL, ktorý dodáva apoproteíny C-II a E pre KM, a od aktivity prenosu apoC-II a apoE do KM. Genetické defekty ktoréhokoľvek z proteínov podieľajúcich sa na metabolizme KM vedú k rozvoju familiárnej hyperchylomikroémie – hromadeniu KM v krvi. Ochorenie sa prejavuje v ranom detstve a je charakterizované hepatosplenomegáliou, pankreatitídou a bolesťami brucha. Ako sekundárny príznak sa pozoruje u pacientov s diabetes mellitus, nefrotickým syndrómom, hypotyreózou a tiež pri zneužívaní alkoholu. Liečba: diéta s nízkym obsahom lipidov (do 30 g/deň) a vysokým obsahom sacharidov.
Typ II – familiárna hypercholesterolémia (hyper-b-lipoproteinémia). Tento typ sa delí na 2 podtypy: IIa, charakterizovaný vysokou hladinou LDL v krvi, a IIb so zvýšenými hladinami LDL aj VLDL. Ochorenie je spojené s poruchou príjmu a katabolizmu LDL (defekt bunkových receptorov pre LDL alebo zmeny v štruktúre LDL), sprevádzané zvýšenou biosyntézou cholesterolu, apo-B a LDL. Toto je najzávažnejšia patológia v metabolizme liekov: riziko vzniku ochorenia koronárnych artérií u pacientov s týmto typom poruchy sa zvyšuje 10-20 krát v porovnaní so zdravými jedincami. Ako sekundárny jav sa môže vyvinúť hyperlipoproteinémia typu II s hypotyreózou a nefrotickým syndrómom. Liečba: Diéta s nízkym obsahom cholesterolu a nasýtených tukov.
Typ III – dys-b-lipoproteinémia (širokopásmová betalipoproteinémia) je spôsobená abnormálnym zložením VLDL. Sú obohatené o voľný cholesterol a defektný apo-E, ktorý inhibuje aktivitu pečeňovej TAG lipázy. To vedie k poruchám v katabolizme cholesterolu a VLDL. Ochorenie sa prejavuje vo veku 30-50 rokov. Stav je charakterizovaný vysokým obsahom zvyškov VLDL, pozorujú sa hypercholesterolémia a triacylglycerolémia, xantómy, aterosklerotické lézie periférnych a koronárnych ciev. Liečba: diétna terapia zameraná na chudnutie.
Typ IV – hyperpre-b-lipoproteinémia (hypertriacylglycerolémia). Primárny variant je spôsobený znížením aktivity LPL, zvýšenie hladiny TAG v krvnej plazme nastáva v dôsledku frakcie VLDL, akumulácia CM nie je pozorovaná. Vyskytuje sa len u dospelých a je charakterizovaný rozvojom aterosklerózy, najskôr koronárnych, potom periférnych artérií. Ochorenie je často sprevádzané zníženou toleranciou glukózy. Ako sekundárny prejav sa vyskytuje pri pankreatitíde a alkoholizme. Liečba: diétna terapia zameraná na chudnutie.
Typ V – hyperpre-b-lipoproteinémia s hyperchylomikronémiou. Pri tomto type patológie sú zmeny v krvných lipidových frakciách zložité: zvyšuje sa obsah cholesterolu a VLDL, znižuje sa závažnosť frakcií LDL a HDL. Pacienti majú často nadváhu, môže sa vyvinúť hepatosplenomegália a pankreatitída, ateroskleróza sa nevyvinie vo všetkých prípadoch. Ako sekundárny jav možno hyperlipoproteinémiu V. typu pozorovať pri inzulín-dependentnom diabetes mellitus, hypotyreóze, pankreatitíde, alkoholizme a glykogenóze I. typu. Liečba: diétna terapia zameraná na chudnutie, diéta s nízkym obsahom sacharidov a tukov.
Po absorpcii do črevného epitelu voľné mastné kyseliny a 2-monoglyceridy opäť tvoria triglyceridy a spolu s fosfolipidmi a cholesterolom sú zahrnuté do chylomikrónov. Chylomikróny sú transportované lymfatickým tokom cez hrudný kanál do hornej dutej žily, čím sa dostávajú do celkového krvného obehu.
Vo vnútri chylomikrónu triglyceridy hydrolyzovaný lipoproteínovou lipázou, čo vedie k uvoľňovaniu mastných kyselín na povrch krvných kapilár v tkanivách. To spôsobuje transport mastných kyselín do tkanív a následnú tvorbu chylomikrónových zvyškov ochudobnených o triglyceridy. Tieto zvyšky potom prijímajú estery cholesterolu z lipoproteínov s vysokou hustotou a častice sú rýchlo absorbované pečeňou. Tento transportný systém pre mastné kyseliny získané z potravy sa nazýva exogénny transportný systém.
Je tu tiež endogénny transportný systém, určený na intraorgánový transport mastných kyselín vytvorených v samotnom organizme. Lipidy sú transportované z pečene do periférnych tkanív a späť a sú tiež prenášané z tukových zásob do rôznych orgánov. Transport lipidov z pečene do periférnych tkanív zahŕňa spoločné pôsobenie VLDL, lipoproteínu so strednou hustotou (IDL), lipoproteínu s nízkou hustotou (LDL) a lipoproteínu s vysokou hustotou (HDL). Častice VLDL, podobne ako chylomikróny, pozostávajú z veľkého hydrofóbneho jadra tvoreného triglyceridmi a estermi cholesterolu a povrchovej lipidovej vrstvy pozostávajúcej hlavne z fosfolipidov a cholesterolu.
VLDL sú syntetizované v pečeni a ich hlavnou funkciou je ukladanie tuku v periférnych tkanivách. Akonáhle sa VLDL dostane do krvného obehu, je vystavený pôsobeniu lipoproteínovej lipázy, ktorá hydrolyzuje triglyceridy na voľné mastné kyseliny. Voľné mastné kyseliny odvodené od chylomikrónov alebo VLDL môžu byť použité ako zdroje energie, štrukturálne zložky fosfolipidových membrán alebo premenené späť na triglyceridy a ako také uložené. Chylomikrónové triglyceridy a VLDL sú tiež hydrolyzované pečeňovou lipázou.
Častice VLDL hydrolýzou triglyceridov sa premieňajú na hustejšie, menšie zvyšky bohaté na cholesterol a triglyceridy (LCR), ktoré sú z plazmy odstránené pečeňovými lipoproteínovými receptormi alebo môžu byť premenené na LDL. LDL je hlavným lipoproteínovým nosičom cholesterolu.
Návrat z periférnych tkanív do pečene sa často nazýva reverzný transport cholesterolu. Častice HDL sa podieľajú na tomto procese odoberaním cholesterolu z tkanív a iných lipoproteínov a jeho transportom do pečene na následné vylúčenie. Ďalším typom transportu, ktorý existuje medzi orgánmi, je prenos mastných kyselín z tukových zásob do orgánov na oxidáciu.
Mastné kyseliny, získané najmä v dôsledku hydrolýzy triglyceridov v tukovom tkanive, sa vylučujú do plazmy, kde sa spájajú s albumínom. Mastné kyseliny viazané na albumín sa koncentračným gradientom transportujú do tkanív s aktívnym metabolizmom, kde sa využívajú predovšetkým ako zdroje energie.
Za posledných 20 rokov len zopár výskumu boli venované problematike transportu lipidov v perinatálnom období (výsledky týchto štúdií nie sú prezentované v tejto publikácii). Potreba podrobnejšieho štúdia tohto problému je zrejmá.
Ako stavebné kamene sa používajú mastné kyseliny materiál ako súčasť lipidov bunkovej steny, ako zdroje energie a sú tiež uložené „v rezerve“ vo forme triglyceridov, hlavne v tukovom tkanive. Niektoré omega-6 a omega-3 LCPUFA sú prekurzormi bioaktívnych metabolitov používaných v bunkovej signalizácii, génovej regulácii a iných metabolicky aktívnych systémoch.
Otázka o úlohe LCPUFA ARA a DHA v procese rastu a vývoja dieťaťa boli jednou z najdôležitejších otázok vo výskume realizovanom v oblasti detskej výživy za posledné dve desaťročia.
Lipidy sú jednou z hlavných zložiek bunkových membrán. Významné množstvo výskumov v oblasti fyziológie lipidov sa zameralo na dve mastné kyseliny – ARA a DHA. ARA sa nachádza v bunkových membránach všetkých štruktúr ľudského tela; je prekurzorom eikozanoidov série 2, leukotriénov série 3 a iných metabolitov, ktoré sú zapojené do bunkových signalizačných systémov a procesu génovej regulácie. Výskum DHA často poukazuje na jej štrukturálne a funkčné úlohy v bunkových membránach.
Toto mastné kyseliny nachádza sa vo vysokých koncentráciách v sivej hmote mozgu, ako aj v tyčinkách a čapiciach sietnice. Štúdie o postupnom vyraďovaní omega-3 mastných kyselín u zvierat ukázali, že 22-uhlíkové omega-6 LCPUFA (napr. 22:5 n-6) môžu štrukturálne, ale nie funkčne, nahradiť 22:6 n-3. Pri neadekvátnych hladinách 22:6 n-3 v tkanivách sa zisťujú zrakové a kognitívne poruchy. Ukázalo sa, že zmena hladín 22:6 n-3 tkaniva ovplyvňuje funkciu neurotransmiterov, aktivitu iónových kanálov, signálne dráhy a génovú expresiu.

Späť na obsah sekcie "
Keďže lipidy sú v podstate hydrofóbne molekuly, transportujú sa vo vodnej fáze krvi ako súčasť špeciálnych častíc – lipoproteínov.
Štruktúru transportných lipoproteínov možno porovnať s orech kto má škrupina A jadro. „Plášť“ lipoproteínu je hydrofilný, jadro je hydrofóbne.
- vzniká povrchová hydrofilná vrstva fosfolipidy(ich polárna časť), cholesterolu(jeho skupina OH), veveričky. Hydrofilita lipidov povrchovej vrstvy je navrhnutá tak, aby zabezpečila rozpustnosť lipoproteínovej častice v krvnej plazme,
- "jadro" je tvorené nepolárnym estery cholesterolu(HS) a triacylglyceroly(TAG), čo sú transportované tuky. Ich pomer sa líši v rôznych typoch lipoproteínov. Smerom do stredu sú tiež zvyšky mastných kyselín fosfolipidov a cyklická časť cholesterolu.
Schéma štruktúry akéhokoľvek transportného lipoproteínu
Existujú štyri hlavné triedy lipoproteínov:
- lipoproteíny s vysokou hustotou (HDL, α-lipoproteíny, α-LP),
- lipoproteíny s nízkou hustotou (LDL, β-lipoproteíny, β-LP),
- lipoproteíny s veľmi nízkou hustotou (VLDL, pre-β-lipoproteíny, pre-β-LP),
- chylomikróny (CM).
Vlastnosti a funkcie lipoproteínov rôznych tried závisia od ich zloženia, t.j. na type prítomných bielkovín a na pomere triacylglycerolov, cholesterolu a jeho esterov, fosfolipidov.

Porovnanie veľkosti a vlastností lipoproteínov
Funkcie lipoproteínov
Funkcie krvných lipoproteínov sú
1. Prenos do buniek tkanív a orgánov
- nasýtené a mononenasýtené mastné kyseliny v zložení triacylglycerolov na následné skladovanie alebo použitie ako energetické substráty,
- polynenasýtené mastné kyseliny v esteroch cholesterolu na použitie bunkami pri syntéze fosfolipidov alebo tvorbe eikosanoidov,
- cholesterol ako membránový materiál,
- fosfolipidy ako membránový materiál,
Za transport sú primárne zodpovedné chylomikróny a VLDL mastné kyseliny ako súčasť TAG. Lipoproteíny s vysokou a nízkou hustotou - pre transport voľných cholesterolu A mastné kyseliny ako súčasť svojich éterov. HDL je tiež schopný darovať bunkám časť svojej fosfolipidovej membrány.
2. Odstránenie prebytočného cholesterolu z bunkových membrán.
3. Transport vitamínov rozpustných v tukoch.
4. Prenos steroidných hormónov (spolu so špecifickými transportnými proteínmi).
Lipoproteínové apoproteíny
Proteíny v lipoproteínoch sa zvyčajne nazývajú apowhites, existuje ich niekoľko typov – A, B, C, D, E. V každej triede lipoproteínov existujú zodpovedajúce apoproteíny, ktoré plnia svoju vlastnú funkciu:
1. Štrukturálne funkcia (" stacionárne» proteíny) – viažu lipidy a tvoria proteín-lipidové komplexy:
- apoB-48– pridáva triacylceroly,
- apoB-100- viaže triacylglyceroly a estery cholesterolu,
- apoA-I- prijíma fosfolipidy,
- apoA-IV- viaže sa na cholesterol.
2. Kofaktor funkcia (" dynamický» proteíny) – ovplyvňujú aktivitu lipoproteínových metabolických enzýmov v krvi.