Dominanța incompletă este un tip special de interacțiune a alelelor genice în care o trăsătură recesivă mai slabă nu poate fi complet suprimată de una dominantă. În conformitate cu legile descoperite de G. Mendel, suprimă complet manifestarea recesivului. Cercetătorul a studiat trăsăturile contrastante pronunțate la plante cu manifestarea fie alelelor dominante, fie recesive. În unele cazuri, Mendel s-a confruntat cu eșecul acestui tipar, dar nu l-a explicat.
Noua formă de moștenire
Uneori, ca urmare a încrucișării, descendenții au moștenit caracteristici intermediare pe care gena părinte nu le-a produs în formă homozigotă. Dominanța incompletă nu a apărut în aparatul conceptual al geneticii până la începutul secolului al XX-lea, când legile lui Mendel au fost redescoperite. În același timp, mulți naturaliști au efectuat experimente genetice cu obiecte vegetale și animale (roșii, leguminoase, hamsteri, șoareci, muște de fructe).
După confirmarea citologică în 1902 de către Walter Setton a legilor mendeliane, principiile transmiterii și interacțiunii trăsăturilor au început să fie explicate din perspectiva comportamentului cromozomilor într-o celulă.
În același 1902, Chermak Correns a descris un caz când, după încrucișarea plantelor cu corolele albe și roșii, descendenții aveau flori roz - dominație incompletă. Această manifestare apare la hibrizi (genotip Ahh) o trăsătură care este intermediară în raport cu dominanta homozigotă ( AA) și recesive ( ahh) fenotipuri. Un efect similar a fost descris pentru multe tipuri de plante cu flori: snapdragon, zambile, frumusețea nopții, căpșuni.
Este dominanța incompletă motivul modificărilor funcționării enzimelor?
Mecanismul de apariție a celei de-a treia variante a trăsăturii poate fi explicat din punctul de vedere al activității enzimelor, care sunt proteine prin natură, iar genele determină structura proteinei. Într-o plantă cu genotip dominant homozigot ( AA) vor fi suficiente enzime, iar cantitatea de pigment va fi normală pentru a colora intens seva celulară.
La homozigoți cu alele recesive ale genei ( ahh) sinteza pigmentului este afectată, corola rămâne necolorată. În cazul unui genotip heterozigot intermediar ( Ahh), gena dominantă oferă încă o cantitate din enzima responsabilă de pigmentare, dar nu este suficientă pentru o culoare strălucitoare, saturată. Rezultatul este o „jumătate” de culoare.
Trăsături moștenite după tipul intermediar
O astfel de moștenire incompletă este bine monitorizată pentru trăsături cu manifestare variabilă:
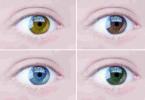
- Intensitatea culorii. W. Batson, încrucișând găini andaluze albe și negre, a obținut urmași cu penaj argintiu. Acest mecanism este prezent și în determinarea culorii irisului ochilor unei persoane.
- Gradul de exprimare a trăsăturii. Structura părului uman este determinată și de moștenirea incompletă a trăsăturii. Genotip AA dă păr creț, ahh- drepte, iar persoanele cu ambele alele au părul ondulat.
- Indicatori măsurabili. Lungimea spicului de grâu se moștenește după principiul dominației incomplete.

În generația F2, numărul de fenotipuri coincide cu numărul de genotipuri, ceea ce caracterizează dominanța incompletă. Încrucișările de testare nu sunt necesare pentru a determina hibrizii, deoarece acestea diferă ca aspect de linia pură dominantă.
Împărțirea caracterelor în timpul traversării
Dominanța completă și incompletă pe măsură ce interacțiunea genelor are loc în conformitate cu aritmetica legilor lui G. Mendel. În primul caz, raportul dintre fenotipuri în F2 (3:1) nu coincide cu raportul dintre genotipurile descendenților (1:2:1), deoarece fenotipic combinațiile de alele AAȘi Ahh apar la fel. Apoi dominanța incompletă este coincidența în F2 a proporției diferitelor genotipuri și fenotipuri (1:2:1).
La căpșuni, culoarea se moștenește an de an după principiul dominanței incomplete. Dacă încrucișați o plantă cu fructe de pădure roșii ( AA) și o plantă cu boabe albe - genotip ahh, apoi în prima generație toate plantele rezultate vor produce fructe cu o culoare roz ( Ahh).

Încrucișând hibrizi din F1, în a doua generație F2 obținem raportul descendenților care coincide cu cel al genotipurilor: 1AA+2Aa+1aa. 25% dintre plantele din a doua generație vor produce fructe roșii și necolorate, 50% dintre plante vor produce fructe roz.
Vom observa o imagine similară în două generații când încrucișăm linii pure de flori de frumusețe nocturnă cu corolele de flori violet și albe.

Particularități ale moștenirii în caz de letalitate genetică
În unele cazuri, este dificil să se determine modul în care genele interacționează din corelarea fenotipurilor descendenților. În a doua generație, împărțirea cu dominație incompletă diferă de cea așteptată 1:2:1 și de 3:1 în dominanță completă. Acest lucru se întâmplă atunci când o trăsătură dominantă sau recesivă într-o stare homozigotă dă un fenotip care este incompatibil cu viața (gene letale).
La oile gri Karakul, mieii nou-născuți homozigoți pentru alela de culoare dominantă mor din cauza faptului că acest genotip provoacă tulburări în dezvoltarea stomacului.
La om, un exemplu de letalitate a formei dominante a genei este brahidactilia (cu degetele scurte). Trăsătura este detectată în cazul unui genotip heterozigot, în timp ce homozigotele dominante mor în stadiile incipiente ale dezvoltării intrauterine.

Alelele genelor recesive pot fi, de asemenea, letale. Anemia falciforme duce, în cazul apariției a două alele recesive în genotip, la o modificare a formei globulelor roșii. Celulele sanguine nu pot absorbi eficient oxigenul, iar 95% dintre copiii cu această anomalie mor din cauza lipsei de oxigen. La heterozigoți, forma modificată a eritrocitelor nu afectează în aceeași măsură viabilitatea.
Divizarea trăsăturilor în prezența genelor letale
În prima generație la trecere AA x aa letalitatea nu se va manifesta, deoarece toți descendenții vor avea genotipul Ah. Iată exemple de împărțire a caracterelor în a doua generație pentru cazurile cu gene letale:
Este important de înțeles că ambele alele acționează în caz de dominanță incompletă, iar efectul suprimării parțiale a trăsăturii este rezultatul interacțiunii produselor genetice.
Interacțiunea genelor alelice sau interacțiune intra-alelică:
A) dominație completă- aceasta este moștenirea în care o genă dominantă o suprimă complet pe una recesivă; în acest tip de dominanță, alelele dominante (genele) își manifestă efectul atât în stare homozigotă, cât și în cea heterozigotă, iar alelele recesive se manifestă fenotipic doar în stare homozigotă.
Homo- și heterozigoții nu se pot distinge;
b) dominație incompletă- aceasta este moștenirea în care gena dominantă nu o suprimă complet pe cea recesivă, în acest caz, hibrizii din prima generație au o trăsătură intermediară, adică există o natură intermediară a moștenirii, de exemplu, moștenirea culorii florii într-un frumusețea nopții sau penajul albastru la găini, culoarea blanii la o vite mare etc.
Homo- și heterozigoții diferă unul de celălalt.
Trăsăturile nu pot fi întotdeauna clar împărțite în dominante și recesive. În aceste cazuri, gena dominantă nu suprimă complet gena recesivă din perechea alelică. În acest caz, vor apărea caracteristici intermediare, iar trăsătura la indivizii homozigoți nu va fi aceeași ca la indivizii heterozigoți. Acest fenomen se numește dominanță incompletă. Să explicăm cu un exemplu.
Când încrucișați o plantă de frumusețe nocturnă care are flori violet (AA) cu o plantă care are flori albe (aa), toate plantele din prima generație vor avea o culoare roz intermediară (Figura 51). Acest lucru nu contrazice regula de uniformitate a hibrizilor din prima generație a lui G. Mendel: într-adevăr, în prima generație toate florile sunt roz. Când se încrucișează doi indivizi de o frumusețe nocturnă din prima generație, despărțirea are loc în a doua generație, dar nu într-un raport de 3: 1, ci într-un raport de 1: 2: 1, adică o floare albă (aa), două roz (Aa) și unul violet (AA).
Până acum, am luat în considerare cazul moștenirii unei trăsături determinate de o genă. Dar orice organism are un număr mare de semne, iar acestea nu sunt doar caracteristici externe, vizibile, ci și semne biochimice (structura moleculelor, activitatea enzimatică, concentrația de substanțe în țesuturile sanguine etc.), anatomice (dimensiunea și forma organelor). ), etc n. Orice semn, oricât de simplu ar părea, depinde de numeroase procese fiziologice și biochimice, fiecare dintre acestea, la rândul lor, depinde de activitatea proteinelor enzimatice.
Totalitatea tuturor caracteristicilor și proprietăților externe și interne ale unui organism se numește fenotip.
Totalitatea tuturor genelor unui organism se numește genotip.
Moștenirea culorii florii în frumusețea nopții:
R: AA × aa
roșu alb
G: Da
F 1: Aa
Trandafir.
P (F 1): Aa × Aa
Trandafir. trandafiri
G: A a A a
F 2: AA Aa Aa aa
Kras. trandafiri trandafiri alb
Dominanță incompletă. Universalitatea legii dominației a fost pusă sub semnul întrebării la scurt timp după redescoperirea ei pe baza unui număr de fapte. S-a dovedit că un număr mare de trăsături la plante și animale sunt caracterizate de moștenire intermediară sau dominație incompletă, in prima generatie. Mendel însuși a remarcat o astfel de moștenire pentru unele trăsături la mazăre.
Cu dominanță incompletă, hibridul F 1 (Aa) nu reproduce în totalitate niciunul dintre caracterele parentale, expresia trăsăturii se dovedește a fi intermediară, cu o prejudecată mai mare sau mai mică către o stare dominantă sau recesivă, dar toți indivizii acestei generație sunt identice ca fenotip.
Un exemplu de dominanță incompletă poate fi culoarea roz intermediară a boabei la hibrizii de căpșuni (Fragaria vesca), obținută din forme de încrucișare cu boabe roșii și albe (Fig. 1).

Figura 1. Moștenirea culorii boabelor cu dominanță incompletă la căpșuni: AA - roșu; aa - alb; La - culoare roz.
Cu dominație completă, divizarea în F 1 după fenotip (3:1) nu coincide cu scindarea după genotip (1:2:1), deoarece heterozigotul Aa nu se poate distinge extern de homozigotul AA. Cu dominanță incompletă în a doua generație, există o coincidență a divizării în fenotip și genotip, deoarece homozigotul dominant AA diferă de heterozigotul Aa. Deci, în exemplul cu căpșuni, împărțirea în F 1 după culoarea boabei este 1 roșu (AA) : 2 roz (Aa) : 1 alb (aa). În acest caz, este posibil doar să se numească culoarea roșie a boabelor dominantă, iar culoarea albă recesivă.
Dominanța incompletă apare și la încrucișarea plantelor de frumusețe nocturnă care au flori violet (AA) cu o plantă care are flori albe (aa). Toate plantele din prima generație vor avea o culoare roz intermediară (Fig. 2). Acest lucru nu contrazice regula de uniformitate a hibrizilor din prima generație a lui G. Mendel: într-adevăr, în prima generație toate florile sunt roz. La încrucișarea a doi indivizi de o frumusețe nocturnă din prima generație, scindarea are loc în a doua generație, dar nu într-un raport de 3:1, ci într-un raport de 1:2:1, adică. o floare este albă (aa), două sunt roz (Aa) și una violet (AA).
Dominanța incompletă s-a dovedit a fi un fenomen larg răspândit, observat în moștenirea culorii florilor la snapdragons, culoarea penajului la pui, culoarea blanii la nurci și cai și multe alte caractere morfologice și fiziologice la plante, animale și oameni.


Figura 2. Fenomenul de dominanță incompletă în timpul formării culorii corolelor florilor de frumusețe nocturnă
Esența dominației. Dominanța este proprietatea unei gene de a determina dezvoltarea unei trăsături într-o stare heterozigotă. Studiile biochimice ale acțiunii genelor, de exemplu la snapdragon (Antirrinum maijus), în care culoarea florii roșii este dominantă față de roșu deschis, demonstrează, de asemenea, natura dominanței. Analiza biochimică a arătat că pigmentul roșu cianidina diferă de pelargonidinul roșu deschis prin hidroxidul de OH suplimentar în poziția a treia (Fig. 3). Fiecare dintre acești doi pigmenți este sintetizat de plantă sub acțiunea unei enzime specifice: unul dintre ei adaugă OH în poziția a 4-a, datorită căreia se produce pelargonidină, iar a doua enzimă o transformă în cianidină, adăugând OH în poziția a 3-a. , adică într-un heterozigot Plantele sintetizează în cele din urmă cianidina, determinând culoarea roșie să domine asupra culorii roșii deschise. Evident, se poate presupune că în toate cazurile în care are loc un mecanism biochimic similar pentru formarea pigmenților, culoarea roșie va domina asupra celei roșii deschise. Într-adevăr, la toate plantele în care se găsește acest mecanism, se observă un model similar de dominanță (streptocarpus, lalea etc.). Analiza biochimică a produselor genetice face posibilă prezicerea naturii dominanței.
 Figura 3. Culoarea florii Snapdragon: AA - roșu;
Figura 3. Culoarea florii Snapdragon: AA - roșu;
aa - roșu deschis și formule chimice ale pigmenților
În cazurile de dominanță incompletă, gena dominantă nu suprimă complet gena recesivă din perechea alelică. În acest caz, vor apărea caracteristici intermediare, iar trăsătura la indivizii homozigoți nu va fi aceeași ca la indivizii heterozigoți. După cum sa dovedit, acest lucru nu înseamnă că alela recesivă este complet suprimată și nu funcționează deloc. De exemplu, cu moștenirea intermediară, ambele alele ale unei gene funcționează. Chiar și în cazul dominanței complete, pot fi găsite exemple care indică faptul că alela recesivă produce un anumit produs. Astfel, in porumb (Zea mays) se gasesc doua tipuri de esteri, care sunt hidrolizati de enzime numite esteraze. Prezența acestor enzime este determinată de o pereche de alele. La plantele heterozigote, doar unul dintre esteri este hidrolizat sub acţiunea unei enzime determinate de alela dominantă. Analiza biochimică a arătat că hibridul produce ambele enzime, dar numai alela dominantă produce o enzimă activă, iar alela recesivă una inactivă. Alela recesivă funcționează în mod egal cu cea dominantă. Este posibil ca în urma interacțiunii acestor două enzime să fie suprimată funcționarea uneia dintre ele, produsă de alela recesivă. Dominanța se manifestă în interacțiunea acelor produse ale acțiunii genelor - trăsături care sunt determinate de alele dominante și recesive.
Managementul dominantei. Mecanismul fenomenului de dominanță determină posibilitatea schimbării dominantei. Se modifică sub influența condițiilor externe, care în diferite grade favorizează sau împiedică dezvoltarea uneia dintr-o pereche de caracteristici. Și într-adevăr, de exemplu, la grâu (Triticum), în condiții normale domină spicul normal, iar în orele scurte de lumină domină cel ramificat. Dar o schimbare a naturii dominanței în dezvoltarea individuală a unui hibrid nu duce la o schimbare a genotipului său și, prin urmare, nu modifică segregarea descendenților săi. Prin urmare, scindarea în F 2 în funcție de forma urechii este aceeași, indiferent de care trăsătură a fost dominantă la hibridul F 1 (Fig. 4).
Figura 4. Dominanța tipului de spic la grâu în funcție de durata orelor de lumină: A - spic normal; a - ramificat.
Pe baza multor ani de cercetare, I.V. Michurin a arătat posibilitatea de a controla dominația la unii hibrizi. Pentru a obține hibrizi cu proprietățile dorite, nu numai că a selectat cu grijă forme pentru încrucișare, dar a creat și condiții favorabile dezvoltării hibridului într-o anumită direcție dorită. Pentru a controla dominația el a dezvoltat o metodă de mentor(educator), care constă în altoirea unui hibrid pe una dintre formele încrucișate. Planta pe care se face altoia se numeste portaltoi, partea altoita a plantei se numeste ram. Atunci când țesuturile vegetale de diferite soiuri sau chiar specii și genuri cresc împreună sub influența portaltoiului, unele caracteristici fiziologice și morfologice se pot schimba în descendent. De exemplu, la polenizarea florilor cireșului Vladimir cu fructe roșii (Prunus cerasus) cu polen din cireșul Winkler cu fructe albe (P. avium), Michurin a obținut un hibrid cu fructe roz pal. O tăietură a unei plante hibride, altoită în coroana unui cireș, a început să producă fructe roz închis. În consecință, sub influența vaccinării, natura dominantei s-a schimbat. Ulterior, prin înmulțirea vegetativă a acestui hibrid s-a obținut soiul Krasa Severa. În selecția pomilor fructiferi care se reproduc vegetativ și, prin urmare, nu își pierd proprietățile dobândite în timpul ontogenezei în timpul reproducerii, metoda mentorului este de mare importanță.
Din păcate, aceste experimente ale lui I.V. Michurin nu au fost însoțite de analize citofiziologice și biochimice, iar esența biochimică a relației dintre componentele grefei nu a fost încă clarificată.
Studiul moștenirii diferitelor caractere pe diferite obiecte a arătat caracterul comun al modelelor de bază de moștenire nu numai pentru plante, ci și pentru animale - vertebrate și nevertebrate, precum și pentru oameni și microorganisme.
Totuși, ca orice lege a naturii vii, legea scindarii se manifestă numai în anumite condiții. Dacă aceste condiții nu sunt îndeplinite, natura divizării se poate modifica.
Cu dominanță incompletă, heterozigoții au un fenotip intermediar între fenotipurile unui homozigot dominant și recesiv. De exemplu, atunci când liniile pure de snapdragons și multe alte specii de plante cu flori cu flori violet și albe sunt încrucișate, indivizii din prima generație au flori roz. Când liniile pure de pui andaluzi alb și negru sunt încrucișate, puii gri se nasc în prima generație. La nivel molecular, cea mai simplă explicație pentru dominanța incompletă poate fi doar o scădere de două ori a activității unei enzime sau a unei alte proteine (dacă alela dominantă produce o proteină funcțională, iar alela recesivă produce una defectuoasă). De exemplu, o alelă defectuoasă care produce o enzimă inactivă poate fi responsabilă pentru colorarea albă, iar o alelă normală care produce o enzimă care produce pigment roșu poate fi responsabilă pentru colorarea roșie. La jumătate din activitatea acestei enzime la heterozigoți, cantitatea de pigment roșu este redusă la jumătate, iar culoarea este roz. Pot exista și alte mecanisme de dominanță incompletă.
Uneori, împărțirea caracterelor din a doua generație se poate abate de la rezultatele așteptate (3:1 - cu dominație completă, 1:2:1 - cu dominanță incompletă). Acest lucru se datorează faptului că, în unele cazuri, homozigoții pentru una dintre trăsături nu sunt viabile. În acest caz, vorbim despre gene letale. O genă poate influența alte trăsături, ducând la scăderea performanței. De exemplu, oile cenușii Karakul, homozigote pentru trăsătura dominantă a colorării cenușii, mor după naștere din cauza subdezvoltării stomacului. Un alt exemplu de genă letală dominantă este brahidactilia umană (degete scurte). Omozigoții pentru această genă mor în stadiile incipiente ale dezvoltării embrionului, iar trăsătura apare doar la heterozigoți.
Un exemplu de genă letală recesivă este gena anemiei falciforme la om. În mod normal, globulele roșii umane au forma unui disc biconcav. Cu anemia secerată, acestea capătă aspectul unei seceri, iar efectul fiziologic se exprimă prin anemie acută și scăderea cantității de oxigen transportată de sânge. La heterozigoți, boala nu se manifestă; totuși, globulele roșii au încă o formă alterată. Homozigoții pentru această trăsătură în 95% din cazuri mor la o vârstă fragedă din cauza deficienței de oxigen, în timp ce heterozigoții sunt destul de viabili.
Când indivizii F1 sunt încrucișați unul cu celălalt, raportul fenotipurilor din F2 diferă de raportul mendelian 3:1 tipic unei încrucișări monohibride. În acest caz, raportul este de 1:2:1, unde jumătate dintre indivizii F2 vor avea același genotip ca F1. Raportul 1: 2: 1 este tipic pentru rezultatele încrucișărilor cu dominanță incompletă.
32. Genetica sexului. Moștenirea legată de sex
Genetica sexului.
Sexul este un ansamblu de caracteristici prin care se realizează o divizare specifică a indivizilor sau celulelor, pe baza unor caracteristici morfologice și fiziologice, care permite îmbinarea înclinațiilor ereditare ale părinților la descendenți în timpul procesului de reproducere sexuală.
Caracteristicile morfologice și fiziologice prin care se realizează o divizare specifică a indivizilor se numesc sexuale.
Caracteristicile asociate cu formarea și funcționarea celulelor germinale sunt numite caracteristici sexuale primare. Acestea sunt gonadele (ovare sau testicule), canalele lor excretoare, glandele accesorii ale aparatului reproducător și organele copulatoare. Toate celelalte caracteristici prin care un sex diferă de celălalt se numesc caracteristici sexuale secundare. Acestea includ: natura liniei părului, prezența și dezvoltarea glandelor mamare, structura scheletului, tipul de dezvoltare a țesutului adipos subcutanat, structura oaselor tubulare etc.
Moștenirea legată de sex- mostenirea unei gene situate pe cromozomii sexuali. Moștenirea trăsăturilor care apar numai la indivizii de un singur sex, dar nu sunt determinate de gene localizate pe cromozomii sexuali, se numește moștenire limitată de sex.
Moștenirea legată de X este moștenirea genelor în cazul în care sexul masculin este heterogametic și se caracterizează prin prezența unui cromozom Y (XY), iar femelele sunt homogametice și au doi cromozomi X (XX). Toate mamiferele (inclusiv oamenii), majoritatea insectelor și reptilelor au acest tip de moștenire.
Moștenirea legată de Z este moștenirea genelor în cazul în care sexul feminin este heterogametic și se caracterizează prin prezența unui cromozom Z (ZW), iar bărbații sunt homogametici și au doi cromozomi Z (ZZ). Toți reprezentanții clasei de păsări au acest tip de moștenire.
Dacă alela unei gene legate de sex situată pe cromozomul X sau pe cromozomul Z este recesivă, atunci trăsătura determinată de această genă se manifestă la toți indivizii de sex heterogametic care au primit această alelă împreună cu cromozomul sexual și la indivizii homozigoți. pentru această alele sex homogametic. Acest lucru se explică prin faptul că al doilea cromozom sexual (Y sau W) din sexul heterogametic nu poartă alele pentru majoritatea sau pentru toate genele găsite pe cromozomul pereche.
Indivizii de sex heterogametic vor avea această trăsătură mult mai des. Prin urmare, bolile care sunt cauzate de alele recesive ale genelor legate de sex afectează bărbații mult mai des, iar femeile sunt adesea purtătoare de astfel de alele.
Observarea între genele alelice; heterozigoții prezintă caracteristici intermediare cu cele ale părinților. În cazul dominanței incomplete la descendenții hibrizilor (F2), împărțirea după genotip și fenotip coincide (1:2:1)
Exemplu: colorarea florilor de frumusețe nocturnă.
O boală genetică umană este brahidactilia - scurtarea falangei mijlocii a degetelor. Gena „b” este responsabilă de dezvoltarea bolii; o persoană sănătoasă are genotipul „BB”. Pacienții cu brahidactilie sunt heterozigoți și au genotipul „BB”. Omozigoții pentru gena „bb” mor înainte de naștere, adică gena „b” în stare homozigotă este letală.
Codominanța– manifestare independentă a genelor alelice. Fiecare genotip prezintă propriul efect și, ca rezultat, prezintă o nouă trăsătură.
Exemplu: formarea celui de-al patrulea grup sanguin la om. Moștenirea grupului de sânge al unei persoane conform sistemului ABO este determinată de gena I; există 3 alele ale acestei gene. I O; IN ABSENTA; I B. Genele I A și I B sunt dominante în raport cu I O, dar codominante între ele, iar la indivizii cu genotipul I A I B apare grupa sanguină 4.
Complementaritatea b – manifestarea unui semn care apare numai în prezența a două gene non-alelice dominante în genotipul organismului.
Exemplu: interacțiunea complementară a observațiilor în moștenirea formelor de pieptene la pui.
 Epistazis– interacțiunea genelor non-alelice, în care gena unei perechi alelice suprimă efectul altei perechi alele. O genă care suprimă acțiunea unei alte gene se numește epistatic
, supresor sau inhibitor. Epistazisul poate fi dominant sau recesiv.
Epistazis– interacțiunea genelor non-alelice, în care gena unei perechi alelice suprimă efectul altei perechi alele. O genă care suprimă acțiunea unei alte gene se numește epistatic
, supresor sau inhibitor. Epistazisul poate fi dominant sau recesiv.
Exemplu: epistaza dominantă: moștenirea definiției culorii la pui. Gena dominantă C – producție normală de pigment; c – nu asigură sinteza enzimatică; gena dominantă I a unei alte perechi alelice este un supresor. Ca urmare, puii cu genotipul C și cu mine se dovedesc a fi albi.
Exemplu: epistasis recesiv este culoarea blanii șoarecilor de casă
Polimerismul– fenomenul de acţiune simultană asupra unei trăsături a mai multor gene non-alelice de acelaşi tip.
Exemplu: moștenirea trăsăturilor cantitative: culoarea semințelor de grâu, pielea umană etc.; producția de ouă, conținutul de zahăr din sfecla de zahăr etc.
Genele sunt responsabile pentru dezvoltarea trăsăturilor cantitative și au un efect cumulativ; cu cât genele sunt mai dominante, cu atât trăsătura este mai strălucitoare.
Exemplu: genele polimerice S 1 S 1 S 2 S 2 sunt responsabile de pigmentarea pielii umane; cu cât genele sunt mai dominante în genotip, cu atât pielea este mai întunecată.
Piele închisă Piele deschisă
R: S 1 S 1 S 2 S 2 x s 1 s 1 s 2 s 2
F1: S 1 s 1 S 2 s 2
Căsătoriile între mulatri vor produce copii cu pigmentare a pielii de la deschis la întuneric. Probabilitatea de a avea un copil cu pielea neagră este de 1/16.
VARIABILITATE.
Variabilitate– capacitatea organismelor fiice de a diferi de formele lor părinte.
Variabilitate
Neereditare Ereditar
(fenotipic, (genotipic,
modificare, individual,
direcțional) aleatoriu)
Mutația combinată
Modificarea variabilității (neereditare) este un răspuns adaptativ al organismului, fixat evolutiv, la schimbările condițiilor de mediu fără modificarea genotipului.
Caracteristic – afectează doar fenotipul; nemoștenit; determinat de condiţiile de existenţă; este de natură adaptabilă la condițiile de mediu.
Modificare(din latinescul „modificatio” - „modificare”) este o modificare neereditară a fenotipului care are loc sub influența factorilor de mediu în limitele normale ale reacției genotipului.
Se numesc limitele în care sunt posibile modificări ale caracteristicilor unui anumit genotip norma de reactie. După gradul de variație a normei de reacție, semnele sunt plastice și neplastice. Plastic (viteză largă de reacție), exemple: conținutul de grăsime al laptelui la vaci, greutatea corpului uman etc. Non-plastic (viteză de reacție îngustă), exemple: culoarea irisului, grupa de sânge uman etc.
Exemple de variabilitate de modificare:
Numărul de globule roșii din 1 ml 3 de sânge aproape se dublează la alpiniștii care se ridică la o înălțime de 4000 m.
Un pin crescut pe un versant de munte va fi foarte diferit ca aspect în ceea ce privește creșterea și forma trunchiului de același pin care crește pe o câmpie.
La oameni, individualitatea și talentul se manifestă ca urmare a influenței înclinațiilor genetice și a mediului extern, iar rolul decisiv în aceasta revine educației și relațiilor sociale.
Variabilitatea modificării este caracterizată de următoarele caracteristici:
ü natura de grup a modificărilor: de exemplu, fiind expuși la razele UV, toți turiștii de pe plajă se bronzează, dar intensitatea bronzului este diferită.
ü adecvarea schimbărilor: cu cât oamenii au fost expuși mai mult la razele soarelui, cu atât au făcut plajă mai mult.
ü durata scurtă a majorității modificărilor: oamenii vin din sud, iar după câteva săptămâni bronzul dispare.
ü adaptabilitatea modificărilor: bronzare – protejarea organismului de efectele razelor UV.
ü limitare: - norma de reactie.
Semnele corpului pot fi:
Calitativ (culoarea ochilor și părului unei persoane);
Cantitativ (înălțimea și greutatea omului).
Pentru a caracteriza gradul de variabilitate al caracteristicilor cantitative se folosește una dintre metodele statistice - construirea unei curbe de variație.
Darwin a numit modificarea schimbărilor definită, deoarece. toți indivizii aceleiași specii, atunci când sunt expuși la condiții similare, se schimbă în același mod, adică o astfel de variabilitate este previzibilă, de exemplu: toate oile, când sunt crescute în condiții mai reci, au început să aibă lână mai groasă.
Genotipic se numește variabilitatea genotipului unui organism.
Caracteristici: mostenit; afectează genotipul; este de natură aleatorie.Se împarte în combinative și mutaționale.
Combinativ variabilitatea este asociată cu obținerea de noi combinații de gene prezente în genotip. Cauzat de: segregarea cromozomală independentă în meioză; combinație aleatorie de cromozomi în timpul fertilizării; recombinarea genelor în timpul încrucișării.
Exemplu: apariția culorii verzi în semințele de mazăre atunci când plantele heterozigote sunt încrucișate cu plante cu semințe galbene.
Variabilitatea mutațională– o modificare a genotipului în sine ca urmare a mutațiilor.
Mutații– modificări bruște și nedirecționate ale ADN-ului, cu apariția în organismele vii a unor semne și proprietăți calitativ noi care nu existau anterior în natură.
Principalele prevederi ale teoriei mutațiilor au fost dezvoltate de G. De Frisone.
Mutațiile apar brusc, ca modificări discrete ale trăsăturilor;
Noile fenotipuri sunt stabile;
Spre deosebire de modificările neereditare, mutațiile nu formează serii continue reprezentând modificări calitative;
Mutațiile pot fi dăunătoare, neutre sau benefice pentru organism;
Probabilitatea de a detecta mutații depinde de numărul de indivizi studiati;
Mutații similare pot apărea în mod repetat.
Cauzele mutațiilor sunt influența factorilor mutaționali de diverse origini. Ele sunt împărțite în:
ü Fizice (radiații ionizante: radiații alfa, beta, gama, raze UV, temperatură ridicată);
ü Chimice (formalină, gaz muștar, medicamente, conservanți alimentari, pesticide etc.);
ü Biologic (virusuri, bacterii).
Mutații genetice asociate cu modificări ale secvenței de nucleotide a ADN-ului unei gene. Tipuri de mutații genetice apar sub formele: duplicări (repetarea unui set de gene localizate în această regiune), inserții, deleții (pierderea secțiunilor de cromozomi în partea de mijloc), inversiuni (rotația unei secțiuni cu 180 o), defiență. (pierderea secțiunilor terminale ale cromozomilor), translocații (transferul unei secțiuni la celălalt capăt al aceluiași cromozom sau la un alt cromozom neomolog).
Exemplu: sfidare - sindromul „strigătul pisicii” - heterozigote pentru defiență pe cromozomul cinci.
Mutații cromozomiale asociat cu mișcarea secțiunilor cromozomilor.
genomic e mutatii– modificarea numărului de cromozomi din genomul unei celule (în cariotipul unui individ).
Genomul– conținutul de material ereditar din setul haploid de cromozomi.
Mutațiile genomice includ:
Poliploidie(euploidie) – o creștere multiplă a numărului haploid de cromozomi.
Celulele cu numere diferite de seturi haploide se numesc triploide (3), tetraploide (4), hexaploide (6) etc.
Poliploizii se formează atunci când divergența cromozomilor la polii celulari în mitoză este întreruptă. Poliploidia este comună în principal la plante. Formele poliploide au frunze, flori, fructe și semințe mai mari. Multe plante cultivate sunt poliploide. Există 2 tipuri de poliploidie: autopoliploidie și alopoliploidie.
Heteroploidie(aneuploidia) este un tip de mutație genomică în care are loc o creștere sau o modificare non-haploidă a numărului de cromozomi. (2n-1 – monosomie, 2n+1 – trisomie; polisomie etc.).
La om, aneuploidia duce la infertilitate și adesea la boli cromozomiale (sindromul Down 2n = 47; sindromul Sherinevsky-Turner, sindromul Klinefelter etc.).
Mutațiile sunt clasificate:
1) din motivele care au provocat mutația: spontane (în condiții naturale) și induse (sub influența vizată a factorilor de mutație asupra organelor). Aceste mutații au fost obținute pentru prima dată de G. A. Nadson și L. S. Filippov (1925) la iradierea ciupercilor cu radiu și de G. Meller (1927) la iradierea muștelor Drosophila cu raze X.
2) prin natura celulelor mutante: somatice - se manifestă în individ însuși, nu sunt moștenite în timpul reproducerii sexuale, ci sunt moștenite în timpul reproducerii vegetative. Exemplu: culori diferite ale irisului la om) și generative - apar în celulele germinale, sunt moștenite și se dezvăluie fenotipic la descendenți, care sunt materialul selecției naturale.
3) în funcție de rezultatul pentru organism: negativ – letal/semi-letal (viabilitate scăzută); neutru; pozitiv (rar).
SELECŢIE
Creșterea (din latinescul „selecție” - „selecție”, „alegere”) este producerea de noi soiuri de plante, rase de animale și tulpini de microorganisme cu proprietăți valoroase pentru oameni.
O rasă, soi sau tulpină este o populație de organisme create artificial de om și caracterizate prin anumite caracteristici ereditare.
Baza teoretică a selecției este genetica.
Principalele metode de selecție sunt:
hibridizare;
poliploidie;
mutageneza;
inginerie celulară și genetică.
N.I.Vavilov - a formulat legea seriei omologice în variabilitatea ereditară; doctrina găsirii de material pentru selecție este ideea pe care a creat-o despre centrele de origine a plantelor cultivate. El a identificat 7 astfel de centre.
I.V. Michurin a avut o contribuție semnificativă la selecția culturilor de fructe. A efectuat metode de hibridizare, selecție și expunere la condițiile de mediu („metoda mentor”) pe dezvoltarea hibrizilor. Un loc important în activitatea de reproducere a lui Michurin a fost ocupat de controlul dominației, care se bazează pe ideea că în condiții specifice de mediu la hibrizi, trăsăturile favorabile acestor condiții primesc o dezvoltare preferențială.