Incomplete dominance is a special type of interaction of gene alleles in which a weaker recessive trait cannot be completely suppressed by a dominant one. In accordance with the laws discovered by G. Mendel, it completely suppresses the manifestation of the recessive. The researcher studied pronounced contrasting traits in plants with the manifestation of either dominant or recessive alleles. In some cases, Mendel was faced with the failure of this pattern, but did not explain it.
New form of inheritance
Sometimes, as a result of crossing, the descendants inherited intermediate characteristics that the parent gene did not produce in homozygous form. Incomplete dominance did not appear in the conceptual apparatus of genetics until the beginning of the 20th century, when Mendel's laws were rediscovered. At the same time, many naturalists conducted genetic experiments with plant and animal objects (tomatoes, legumes, hamsters, mice, fruit flies).
After cytological confirmation in 1902 by Walter Setton of Mendelian laws, the principles of transmission and interaction of traits began to be explained from the perspective of the behavior of chromosomes in a cell.
In the same 1902, Chermak Correns described a case when, after crossing plants with white and red corollas, the descendants had pink flowers - incomplete dominance. This manifestation occurs in hybrids (genotype Ahh) a trait that is intermediate in relation to the homozygous dominant ( AA) and recessive ( ahh) phenotypes. A similar effect has been described for many types of flowering plants: snapdragon, hyacinth, night beauty, strawberries.
Is incomplete dominance the reason for changes in enzyme functioning?
The mechanism for the appearance of the third variant of the trait can be explained from the point of view of the activity of enzymes, which are proteins by nature, and genes determine the structure of the protein. In a plant with a homozygous dominant genotype ( AA) there will be enough enzymes, and the amount of pigment will be normal to intensely color the cell sap.
In homozygotes with recessive alleles of the gene ( ahh) pigment synthesis is impaired, the corolla remains uncolored. In the case of an intermediate heterozygous genotype ( Ahh), the dominant gene still provides some amount of the enzyme responsible for pigmentation, but it is not enough for a bright, saturated color. The result is a “half” color.
Traits inherited by intermediate type
Such incomplete inheritance is well monitored for traits with variable manifestation:
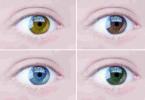
- Color intensity. W. Batson, having crossed black and white Andalusian chickens, obtained offspring with silver plumage. This mechanism is also present in determining the color of the iris of a person’s eyes.
- The degree of expression of the trait. The structure of human hair is also determined by the incomplete inheritance of the trait. Genotype AA gives curly hair, ahh- straight, and people with both alleles have wavy hair.
- Measurable indicators. The length of the wheat ear is inherited according to the principle of incomplete dominance.

In the F2 generation, the number of phenotypes coincides with the number of genotypes, which characterizes incomplete dominance. Test crosses are not required to determine hybrids, since they differ in appearance from the dominant pure line.
Splitting of characters during crossing
Complete and incomplete dominance as gene interaction occurs in accordance with the arithmetic of G. Mendel’s laws. In the first case, the ratio of phenotypes in F2 (3:1) does not coincide with the ratio of genotypes of descendants (1:2:1), since phenotypically the combinations of alleles AA And Ahh appear the same. Then incomplete dominance is the coincidence in F2 of the proportion of different genotypes and phenotypes (1:2:1).
In strawberries, color is inherited year by year according to the principle of incomplete dominance. If you cross a plant with red berries ( AA) and a plant with white berries - genotype ahh, then in the first generation all the resulting plants will produce fruits with a pink color ( Ahh).

Having crossed hybrids from F1, in the second generation F2 we obtain the ratio of descendants that coincides with that of the genotypes: 1AA+2Aa+1aa. 25% of plants from the second generation will produce red and uncolored fruits, 50% of plants will produce pink ones.
We will observe a similar picture in two generations when crossing pure lines of night beauty flowers with corollas of purple and white flowers.

Peculiarities of inheritance in case of gene lethality
In some cases, it is difficult to determine how genes interact from the correlation of offspring phenotypes. In the second generation, splitting with incomplete dominance differs from the expected 1:2:1, and from 3:1 in complete dominance. This happens when a dominant or recessive trait in a homozygous state gives a phenotype that is incompatible with life (lethal genes).
In gray Karakul sheep, newborn lambs homozygous for the dominant color allele die due to the fact that this genotype causes disturbances in the development of the stomach.
In humans, an example of the lethality of the dominant form of the gene is brachydactyly (short-fingered). The trait is detected in the case of a heterozygous genotype, while dominant homozygotes die in the early stages of intrauterine development.

Recessive gene alleles can also be lethal. Sickle cell anemia leads, in the case of the appearance of two recessive alleles in the genotype, to a change in the shape of red blood cells. Blood cells cannot effectively absorb oxygen, and 95% of children with this anomaly die from oxygen starvation. In heterozygotes, the altered shape of erythrocytes does not affect viability to the same extent.
Splitting of traits in the presence of lethal genes
In the first generation when crossing AA x aa lethality will not manifest itself, since all descendants will have the genotype Ah. Here are examples of character splitting in the second generation for cases with lethal genes:
It is important to understand that both alleles act in case of incomplete dominance, and the effect of partial suppression of the trait is the result of the interaction of gene products.
Interaction of allelic genes or intra-allelic interaction:
A) complete dominance- this is inheritance in which a dominant gene completely suppresses a recessive one; in this type of dominance, dominant alleles (genes) manifest their effect in both homozygous and heterozygous states, and recessive alleles are phenotypically manifested only in a homozygous state.
Homo- and heterozygotes are indistinguishable;
b) incomplete dominance- this is inheritance in which the dominant gene does not completely suppress the recessive one, in this case, the first generation hybrids have an intermediate trait, that is, there is an intermediate nature of inheritance, for example, inheritance of flower color in a night beauty, or blue plumage in chickens, coat color in a large cattle, etc.
Homo- and heterozygotes differ from each other.
Traits cannot always be clearly divided into dominant and recessive. In these cases, the dominant gene does not completely suppress the recessive gene from the allelic pair. In this case, intermediate characteristics will arise, and the trait in homozygous individuals will not be the same as in heterozygous individuals. This phenomenon is called incomplete dominance. Let's explain it with an example.
When you cross a night beauty plant that has purple flowers (AA) with a plant that has white flowers (aa), all of the first generation plants will have an intermediate pink color (Figure 51). This does not contradict G. Mendel’s rule of uniformity of hybrids of the first generation: indeed, in the first generation all the flowers are pink. When crossing two individuals of a night beauty from the first generation, splitting occurs in the second generation, but not in a ratio of 3: 1, but in a ratio of 1: 2: 1, i.e. one white flower (aa), two pink (Aa) and one purple (AA).
Until now, we have considered the case of inheritance of one trait determined by one gene. But any organism has a huge number of signs, and these are not only external, visible features, but also biochemical signs (structure of molecules, enzyme activity, concentration of substances in blood tissues, etc.), anatomical (size and shape of organs), etc. n. Any sign, no matter how simple it may seem, depends on numerous physiological and biochemical processes, each of which, in turn, depends on the activity of enzyme proteins.
The totality of all external and internal characteristics and properties of an organism is called a phenotype.
The totality of all the genes of an organism is called a genotype.
Inheritance of flower color in the night beauty:
R: AA × aa
Red white
G: Oh yeah
F 1: Aa
Rose.
P (F 1): Aa × Aa
Rose. roses
G: A a A a
F 2: AA Aa Aa aa
Kras. roses roses white
Incomplete dominance. The universality of the law of dominance was questioned soon after its rediscovery on the basis of a number of facts. It turned out that a large number of traits in plants and animals are characterized by intermediate inheritance, or incomplete dominance, in the first generation. Mendel himself noted such inheritance for some traits in peas.
With incomplete dominance, the hybrid F 1 (Aa) does not fully reproduce any of the parental characters, the expression of the trait turns out to be intermediate, with a greater or lesser bias towards a dominant or recessive state, but all individuals of this generation are identical in phenotype.
An example of incomplete dominance can be the intermediate pink color of the berry in strawberry hybrids (Fragaria vesca), obtained from crossing forms with red and white berries (Fig. 1).

Figure 1. Inheritance of berry color with incomplete dominance in strawberries: AA - red; aa - white; La - pink color.
With complete dominance, the splitting in F 1 by phenotype (3:1) does not coincide with the splitting by genotype (1:2:1), since heterozygote Aa is externally indistinguishable from homozygote AA. With incomplete dominance in the second generation, there is a coincidence of splitting in phenotype and genotype, since the dominant homozygote AA differs from the heterozygote Aa. So, in the example with strawberries, the split in F 1 according to the color of the berry is 1 red (AA) : 2 pink (Aa) : 1 white (aa). In this case, it is only conditionally possible to call the red color of the berry dominant, and the white color recessive.
Incomplete dominance also occurs when crossing night beauty plants that have purple flowers (AA) with a plant that has white flowers (aa). All plants of the first generation will have an intermediate pink color (Fig. 2). This does not contradict G. Mendel’s rule of uniformity of hybrids of the first generation: indeed, in the first generation all the flowers are pink. When crossing two individuals of a night beauty from the first generation, splitting occurs in the second generation, but not in a ratio of 3:1, but in a ratio of 1:2:1, i.e. one flower is white (aa), two are pink (Aa) and one is purple (AA).
Incomplete dominance has proven to be a widespread phenomenon, observed in the inheritance of flower color in snapdragons, plumage color in chickens, coat color in minks and horses, and many other morphological and physiological characters in plants, animals, and humans.


Figure 2. The phenomenon of incomplete dominance during the formation of the color of the corollas of night beauty flowers
The essence of dominance. Dominance is the property of a gene to determine the development of a trait in a heterozygous state. Biochemical studies of gene action, for example in snapdragon (Antirrinum maijus), in which red flower color is dominant over light red, also demonstrate the nature of dominance. Biochemical analysis showed that the red pigment cyanidin differs from the light red pelargonidin by the extra OH hydroxide in the third position (Fig. 3). Each of these two pigments is synthesized by the plant under the action of a specific enzyme: one of them adds OH to the 4th position, due to which pelargonidin is produced, and the second enzyme converts it into cyanidin, adding OH to the 3rd position, i.e. in a heterozygous Plants eventually synthesize cyanidin, causing the red color to dominate over the light red color. Obviously, it can be assumed that in all cases where a similar biochemical mechanism for the formation of pigments takes place, the red color will dominate over the light red one. Indeed, in all plants where this mechanism is found, a similar pattern of dominance is observed (streptocarpus, tulip, etc.). Biochemical analysis of gene products makes it possible to predict the nature of dominance.
 Figure 3. Snapdragon flower color: AA - red;
Figure 3. Snapdragon flower color: AA - red;
aa - light red and chemical formulas of pigments
In cases of incomplete dominance, the dominant gene does not completely suppress the recessive gene from the allelic pair. In this case, intermediate characteristics will arise, and the trait in homozygous individuals will not be the same as in heterozygous individuals. As it turned out, this does not mean that the recessive allele is completely suppressed and does not function at all. For example, with intermediate inheritance, both alleles of a gene function. Even in the case of complete dominance, examples can be found indicating that the recessive allele produces a certain product. Thus, in corn (Zea mays) two types of esters are found, which are hydrolyzed by enzymes called esterases. The presence of these enzymes is determined by a pair of alleles. In heterozygous plants, only one of the esters is hydrolyzed under the action of an enzyme determined by the dominant allele. Biochemical analysis showed that the hybrid produces both enzymes, but only the dominant allele produces an active enzyme, and the recessive allele produces an inactive one. The recessive allele functions on an equal basis with the dominant one. It is possible that as a result of the interaction of these two enzymes, the functioning of one of them, produced by the recessive allele, is suppressed. Dominance is manifested in the interaction of those products of gene action - traits that are determined by dominant and recessive alleles.
Dominance management. The mechanism of the phenomenon of dominance determines the possibility of changing dominance. It changes under the influence of external conditions, which to varying degrees favor or hinder the development of one of a pair of characteristics. And indeed, for example, in wheat (Triticum), under normal conditions the normal ear dominates, and under short daylight hours, the branched one dominates. But a change in the nature of dominance in the individual development of a hybrid does not lead to a change in its genotype and, therefore, does not change the segregation in its offspring. Therefore, the splitting in F 2 according to the shape of the ear is the same, regardless of which trait was dominant in the F 1 hybrid (Fig. 4).
Figure 4. Dominance of ear type in wheat depending on the length of daylight hours: A - normal ear; a - branched.
Based on many years of research, I.V. Michurin showed the possibility of controlling dominance in some hybrids. In order to obtain hybrids with the desired properties, he not only carefully selected forms for crossing, but also created conditions favorable to the development of the hybrid in a certain, desired direction. To control dominance he developed a mentor method(educator), which consists in grafting a hybrid onto one of the crossed forms. The plant on which the graft is made is called the rootstock, the grafted part of the plant is called the scion. When plant tissues of different varieties or even species and genera grow together under the influence of the rootstock, some physiological and morphological features may change in the scion. For example, when pollinating the flowers of the red-fruited Vladimir cherry (Prunus cerasus) with pollen from the white-fruited Winkler cherry (P. avium), Michurin obtained a hybrid with pale pink fruits. A cutting of a hybrid plant, grafted into the crown of a cherry tree, began to produce dark pink fruits. Consequently, under the influence of vaccination, the nature of dominance changed. Subsequently, through vegetative propagation of this hybrid, the Krasa Severa variety was obtained. In the selection of fruit trees that reproduce vegetatively and therefore do not lose the properties acquired during ontogenesis during reproduction, the mentor method is of great importance.
Unfortunately, these experiments by I.V. Michurin were not accompanied by cytophysiological and biochemical analysis, and the biochemical essence of the relationship between the components of the graft has not yet been clarified.
The study of the inheritance of various characters on different objects showed the commonality of the basic patterns of inheritance not only for plants, but also for animals - vertebrates and invertebrates, as well as for humans and microorganisms.
However, like any law of living nature, the law of splitting manifests itself only under certain conditions. If these conditions are not met, the nature of the splitting may change.
With incomplete dominance, heterozygotes have a phenotype intermediate between the phenotypes of a dominant and recessive homozygote. For example, when pure lines of snapdragons and many other species of flowering plants with purple and white flowers are crossed, the first generation individuals have pink flowers. When pure lines of black and white Andalusian chickens are crossed, gray chickens are born in the first generation. At the molecular level, the simplest explanation for incomplete dominance may be just a twofold decrease in the activity of an enzyme or other protein (if the dominant allele produces a functional protein, and the recessive allele produces a defective one). For example, a defective allele that produces an inactive enzyme may be responsible for white coloration, and a normal allele that produces an enzyme that produces red pigment may be responsible for red coloration. At half the activity of this enzyme in heterozygotes, the amount of red pigment is halved, and the color is pink. There may be other mechanisms of incomplete dominance.
Sometimes the splitting of characters in the second generation may deviate from the expected results (3:1 - with complete dominance, 1:2:1 - with incomplete dominance). This is due to the fact that in some cases homozygotes for one of the traits are not viable. In this case, we talk about lethal genes. One gene can influence other traits, resulting in decreased performance. For example, gray Karakul sheep, homozygous for the dominant trait of gray coloration, die after birth due to underdevelopment of the stomach. Another example of a dominant lethal gene is human brachydactyly (short fingers). Homozygotes for this gene die in the early stages of embryo development, and the trait appears only in heterozygotes.
An example of a recessive lethal gene is the sickle cell anemia gene in humans. Normally, human red blood cells have the shape of a biconcave disc. With sickle cell anemia, they take on the appearance of a sickle, and the physiological effect is expressed by acute anemia and a decrease in the amount of oxygen carried by the blood. In heterozygotes, the disease does not manifest itself; however, the red blood cells still have an altered shape. Homozygotes for this trait in 95% of cases die at an early age due to oxygen deficiency, while heterozygotes are quite viable.
When F1 individuals are crossed with each other, the ratio of phenotypes in F2 differs from the Mendelian 3:1 ratio typical of a monohybrid cross. In this case, the ratio is 1:2:1, where half of the F2 individuals will have the same genotype as F1. The ratio 1: 2: 1 is typical for the results of crosses with incomplete dominance.
32. Genetics of sex. Sex-linked inheritance
Genetics of sex.
Sex is a set of characteristics by which a specific division of individuals or cells is made, based on morphological and physiological characteristics, which allows the combination of the hereditary inclinations of the parents in the descendants during the process of sexual reproduction.
The morphological and physiological characteristics by which a specific division of individuals is made is called sexual.
Characteristics associated with the formation and functioning of germ cells are called primary sexual characteristics. These are the gonads (ovaries or testes), their excretory ducts, accessory glands of the reproductive apparatus, and copulatory organs. All other characteristics by which one sex differs from the other are called secondary sexual characteristics. These include: the nature of the hairline, the presence and development of the mammary glands, the structure of the skeleton, the type of development of subcutaneous fatty tissue, the structure of tubular bones, etc.
Sex-linked inheritance- inheritance of a gene located on the sex chromosomes. Inheritance of traits that appear only in individuals of one sex, but are not determined by genes located on the sex chromosomes, is called sex-limited inheritance.
X-linked inheritance is the inheritance of genes in the case where the male sex is heterogametic and is characterized by the presence of a Y chromosome (XY), and females are homogametic and have two X chromosomes (XX). All mammals (including humans), most insects and reptiles have this type of inheritance.
Z-linked inheritance is the inheritance of genes in the case where the female sex is heterogametic and is characterized by the presence of a Z chromosome (ZW), and males are homogametic and have two Z chromosomes (ZZ). All representatives of the class of birds have this type of inheritance.
If the allele of a sex-linked gene located on the X chromosome or Z chromosome is recessive, then the trait determined by this gene is manifested in all individuals of the heterogametic sex that received this allele along with the sex chromosome, and in individuals homozygous for this allele homogametic sex. This is explained by the fact that the second sex chromosome (Y or W) in the heterogametic sex does not carry alleles for most or all of the genes found on the paired chromosome.
Individuals of the heterogametic sex will have this trait much more often. Therefore, diseases that are caused by recessive alleles of sex-linked genes affect men much more often, and women are often carriers of such alleles
Observation between allelic genes; heterozygotes exhibit characteristics intermediate to those of the parents. In case of incomplete dominance in the offspring of hybrids (F2), the splitting by genotype and phenotype coincides (1:2:1)
Example: coloring of night beauty flowers.
A human genetic disease is brachydactyly - shortening of the middle phalanx of the fingers. The “b” gene is responsible for the development of the disease; a healthy person has the “BB” genotype. Patients with brachydactyly are heterozygotes and have the “BB” genotype. Homozygotes for the “bb” gene die before birth, that is, the “b” gene in the homozygous state is lethal.
Codominance– independent manifestation of allelic genes. Each genotype exhibits its own effect, and as a result they exhibit a new trait.
Example: the formation of the fourth blood group in humans. Inheritance of a person's blood group according to the ABO system is determined by gene I; there are 3 alleles of this gene. I O; I A; I B. Genes I A and I B are dominant in relation to I O, but codominant in relation to each other, and in individuals with the genotype I A I B blood group 4 appears.
Complementarity b – manifestation of a sign that occurs only in the presence of two dominant non-allelic genes in the genotype of the organism.
Example: complementary interaction of observations in the inheritance of comb shapes in chickens.
 Epistasis– interaction of non-allelic genes, in which the gene of one allelic pair suppresses the effect of another allelic pair. A gene that suppresses the action of another gene is called epistatic
, suppressor or inhibitor. Epistasis can be dominant or recessive.
Epistasis– interaction of non-allelic genes, in which the gene of one allelic pair suppresses the effect of another allelic pair. A gene that suppresses the action of another gene is called epistatic
, suppressor or inhibitor. Epistasis can be dominant or recessive.
Example: dominant epistasis: inheritance of color definition in chickens. Dominant gene C – normal pigment production; c – does not provide enzyme synthesis; the dominant gene I of another allelic pair is a suppressor. As a result, chickens with genotypes C and I turn out to be white.
Example: recessive epistasis is the coat color of house mice
Polymerism– the phenomenon of simultaneous action on a trait of several non-allelic genes of the same type.
Example: inheritance of quantitative traits: color of wheat seeds, human skin, etc.; egg production, sugar content in sugar beets, etc.
Genes are responsible for the development of quantitative traits and have a cumulative effect; the more dominant genes, the brighter the trait.
Example: polymer genes S 1 S 1 S 2 S 2 are responsible for the pigmentation of human skin; the more dominant genes in the genotype, the darker the skin.
Dark skin Light skin
R: S 1 S 1 S 2 S 2 x s 1 s 1 s 2 s 2
F1: S 1 s 1 S 2 s 2
Marriages between mulattoes will produce children with skin pigmentation ranging from light to dark. The probability of having a child with black skin is 1/16.
VARIABILITY.
Variability– the ability of daughter organisms to differ from their parent forms.
Variability
Non-hereditary Hereditary
(phenotypic, (genotypic,
modification, individual,
directional) random)
Combinative Mutation
Modifying variability (non-hereditary) is an evolutionarily fixed adaptive response of the body to changes in environmental conditions without changing the genotype.
Characteristic – affects only the phenotype; not inherited; determined by the conditions of existence; is adaptive in nature to environmental conditions.
Modification(from the Latin “modificatio” - “modification”) is a non-hereditary change in phenotype that occurs under the influence of environmental factors within the normal limits of the genotype reaction.
The limits within which changes in characteristics of a given genotype are possible are called reaction norm. According to the degree of variation in the reaction norm, signs are plastic and non-plastic. Plastic (wide reaction rate), examples: fat content of milk in cows, human body weight, etc. Non-plastic (narrow reaction rate), examples: color of the iris, human blood type, etc.
Examples of modification variability:
The number of red blood cells in 1 ml 3 of blood almost doubles in climbers who rise to a height of 4000 m.
A pine tree grown on a mountain slope will be very different in appearance in terms of growth and trunk shape from the same pine tree growing on a plain.
In people, individuality and talent are manifested as a result of the influence of genetic inclinations and the external environment, and the decisive role in this belongs to upbringing and social relations.
Modification variability is characterized by the following features:
ü group nature of the changes: for example, being exposed to UV rays, all vacationers on the beach tan, but the intensity of the tan is different.
ü adequacy of changes: the more people were exposed to the rays of the sun, the more they sunbathed.
ü the short duration of most modifications: people come from the south, and after a few weeks the tan goes away.
ü adaptability of changes: tanning – protecting the body from the effects of UV rays.
ü limitation: - reaction norm.
Signs of the body may be:
Qualitative (color of a person’s eyes and hair);
Quantitative (human height and weight).
To characterize the degree of variability of quantitative characteristics, one of the statistical methods is used - constructing a variation curve.
Darwin called the modification of changes definite, because. all individuals of the same species, when exposed to similar conditions, change in the same way, i.e., such variability is predictable, for example: all sheep, when raised in colder conditions, began to have thicker wool.
Genotypic is called the variability of the genotype of an organism.
Characteristics: inherited; affects the genotype; is random in nature. It is divided into combinative and mutational.
Combinative variability is associated with obtaining new combinations of genes present in the genotype. Caused by: independent chromosome segregation in meiosis; random combination of chromosomes during fertilization; recombination of genes during crossing over.
Example: the appearance of green color in pea seeds when heterozygous plants are crossed with plants with yellow seeds.
Mutational variability– a change in the genotype itself as a result of mutations.
Mutations– sudden abrupt and undirected changes in DNA, with the appearance in living organisms of qualitatively new signs and properties that did not previously exist in nature.
The main provisions of mutation theory were developed by G. De Frisone.
Mutations occur suddenly, as discrete changes in traits;
New phenotypes are stable;
Unlike non-hereditary changes, mutations do not form continuous series representing qualitative changes;
Mutations can be harmful, neutral or beneficial for the body;
The probability of detecting mutations depends on the number of individuals studied;
Similar mutations may occur repeatedly.
The causes of mutations are the influence of mutational factors of various origins. They are divided into:
ü Physical (ionizing radiation: alpha, beta, gamma radiation, UV rays, high temperature);
ü Chemical (formalin, mustard gas, drugs, food preservatives, pesticides, etc.);
ü Biological (viruses, bacteria).
Gene mutations associated with changes in the nucleotide sequence of the DNA of one gene. Types of gene mutations appear in the forms: duplications (repetition of a set of genes localized in this region), insertions, deletions (loss of sections of chromosomes in the middle part), inversions (rotation of a section by 180 o), defimency (loss of terminal sections of chromosomes), translocations ( transfer of a section to the other end of the same chromosome or to another non-homologous chromosome).
Example: defimency - “cry of the cat” syndrome - heterozygosity for defimency on the fifth chromosome.
Chromosomal mutations associated with the movement of chromosome sections.
Genomic e mutations– change in the number of chromosomes in the genome of a cell (in the karyotype of an individual).
Genome– the content of hereditary material in the haploid set of chromosomes.
Genomic mutations include:
Polyploidy(euploidy) – a multiple increase in the haploid number of chromosomes.
Cells with different numbers of haploid sets are called triploid (3), tetraploid (4), hexaploid (6), etc.
Polyploids are formed when the divergence of chromosomes to the cell poles in mitosis is disrupted. Polyploidy is common mainly in plants. Polyploid forms have larger leaves, flowers, fruits and seeds. Many cultivated plants are polyploids. There are 2 types of polyploidy: autopolyploidy and allopolyploidy.
Heteroploidy(aneuploidy) is a type of genomic mutation in which a non-haploid increase or change in the number of chromosomes occurs. (2n-1 – monosomy, 2n+1 – trisomy; polysomy, etc.).
In humans, aneuploidy leads to infertility and often to chromosomal diseases (Down syndrome 2n = 47; Sherinevsky-Turner syndrome, Klinefelter syndrome, etc.).
Mutations are classified:
1) for the reasons that caused the mutation: spontaneous (in natural conditions) and induced (under the targeted influence of mutation factors on the organs). These mutations were first obtained by G. A. Nadson and L. S. Filippov (1925) when irradiating fungi with radium, and by G. Meller (1927) when irradiating Drosophila flies with X-rays.
2) by the nature of the mutated cells: somatic - manifested in the individual itself, are not inherited during sexual reproduction, but are inherited during vegetative reproduction. Example: different colors of the iris in humans) and generative - occur in germ cells, are inherited, and are revealed phenotypically in descendants, which are the material for natural selection.
3) according to the outcome for the body: negative – lethal/semi-lethal (decreased viability); neutral; positive (rare).
SELECTION
Breeding (from the Latin “selection” - “selection”, “choice”) is the production of new varieties of plants, animal breeds and strains of microorganisms with properties valuable to humans.
A breed, variety, or strain is a population of organisms artificially created by man and characterized by certain hereditary characteristics.
The theoretical basis of selection is genetics.
The main selection methods are:
hybridization;
polyploidy;
mutagenesis;
cellular and genetic engineering.
N.I. Vavilov - formulated the law of homological series in hereditary variability; the doctrine of finding material for selection is the idea he created about the centers of origin of cultivated plants. He identified 7 such centers.
I.V. Michurin made a significant contribution to the selection of fruit crops. He performed methods of hybridization, selection and exposure to environmental conditions (“mentor method”) on developing hybrids. An important place in Michurin’s breeding work was occupied by the control of dominance, which is based on the idea that under specific environmental conditions in hybrids, traits favorable for these conditions receive preferential development