Обмін речовин, що надходять всередину клітини або виділяються назовні, а також обмін різними сигналами з мікро-і макрооточенням відбувається через зовнішню мембрану клітини. Як відомо, клітинна мембрана є ліпідним біслоєм, в який вбудовані різні білкові молекули, що виконують роль спеціалізованих рецепторів, іонних каналів, пристроїв, які активно переносять або видаляють різні хімічні речовини, міжклітинних контактів і т.п. асиметрично: зовнішня поверхня складається із сфінгомієліну та фосфатидилхоліну, внутрішня - з фосфатидилсерину та фосфатидилетаноламіну. Підтримка такої асиметрії потребує витрат енергії. Тому у разі пошкодження клітини, її інфікування, енергетичного голодування відбувається збагачення зовнішньої поверхні мембрани невластивими їй фосфоліпідами, що стає сигналом для інших клітин та ферментів про пошкодження клітини з відповідною реакцією на це. Найбільшу роль грає розчинна форма фосфоліпази A2, яка розщеплює арахідонову кислоту та створює лізоформи з вищезгаданих фосфоліпідів. Арахидонова кислота є лімітуючою ланкою для створення таких медіаторів запалення, як ейкозаноїди, а до лізоформів в мембрані приєднуються захисні молекули - пентраксини (С-реактивний протеїн (СРП), попередники амілоїдних білків) - з подальшою активацією системи комп'ютера.
Будова мембрани сприяє збереженню особливостей внутрішнього середовища клітини, її відмінностей від зовнішнього середовища. Це забезпечується вибірковою проникністю клітинної мембрани, існуванням у ній механізмів активного транспорту. Їх порушення в результаті прямого пошкодження, наприклад, тетродотоксином, уабаїном, тетраетиламмонием, або у разі недостатнього енергетичного забезпечення відповідних "насосів" призводить до порушення електролітного складу клітини, зміни в ній метаболізму, порушення специфічних функцій - скорочення, проведення імпульсу збудження тощо. Порушення клітинних іонних каналів (кальцієвих, натрієвих, калієвих та хлорних) у людини також може бути генетично обумовлено мутацією генів, які відповідають за структуру цих каналів. Так звані каналопатії спричиняють спадкові хвороби нервової, м'язової, травної систем. Надмірне надходження всередину клітини води може призвести до її розриву – цитолізу – внаслідок перфорації мембрани при активації комплементу або атаки цитотоксичних лімфоцитів та натуральних кілерів.
У клітинну мембрану вбудовано безліч рецепторів – структур, які у разі поєднання з відповідними специфічними сигнальними молекулами (лігандами) передають сигнал усередину клітини. Це відбувається за допомогою різних регуляторних каскадів, що складаються з ферментативно-активних молекул, які послідовно активуються і в кінцевому підсумку сприяють реалізації різних клітинних програм, таких як ріст і проліферація, диференціювання, рухливість, старіння, загибель клітини. Регуляторні каскади досить багато, але їх кількість до кінця ще не визначена. Система рецепторів та об'єднаних з ними регуляторних каскадів існує і всередині клітини; вони створюють певну регуляторну мережу з пунктами концентрації, розподілу та вибору подальшого шляху сигналу залежно від функціонального стану клітини, етапу її розвитку, одночасної дії сигналів з інших рецепторів. Результатом цього може бути гальмування або посилення сигналу, направлення його іншим регуляторним шляхом. Як рецепторний апарат, так і шляхи передачі сигналу через регуляторні каскади, наприклад до ядра, можуть порушуватися в результаті генетичного дефекту, який виникає як вроджена вада на рівні організму або внаслідок соматичної мутації в певному типі клітин. Зазначені механізми можуть пошкоджуватися інфекційними агентами, токсинами, а також змінюватись у процесі старіння. Завершальним етапом може бути порушення функцій клітини, процесів її проліферації і диференціювання.
На поверхні клітин також розташовані молекули, що відіграють важливу роль у процесах міжклітинної взаємодії. До них можуть належати білки клітинної адгезії, антигени тканинної сумісності, тканинноспецифічні, диференціюючі антигени тощо. Зміни у складі цих молекул зумовлюють порушення міжклітинних взаємодій і можуть стати причиною включення відповідних механізмів елімінації таких клітин резервуар інфекції, особливо вірусної, чи як потенційні ініціатори пухлинного зростання.
Порушення енергетичного забезпечення клітини
Джерелом енергії в клітині є продукти харчування, після розщеплення яких до кінцевих речовин виділяється енергія. Головним місцем утворення енергії є мітохондрії, в яких речовини окислюються за допомогою ферментів дихального ланцюга. Окислення - основний постачальник енергії, оскільки в результаті гліколізу з такої кількості субстратів окислення (глюкози) вивільняється, порівняно з окисленням, не більше 5 % енергії. Близько 60 % енергії, що вивільняється при окисленні, акумулюється шляхом окисного фосфорилювання в макроергічних фосфатах (АТФ, креатинфосфат), решта її розсіюється у вигляді тепла. У подальшому макроергічні фосфати використовуються клітиною для таких процесів, як робота насосів, синтез, поділ, рух, секреція та ін. , третій – механізм використання енергії.
Порушення електронного транспорту в дихальному ланцюгу мітохондрій або роз'єднання окислення і фосфорилювання АДФ при втраті протонного потенціалу - рушійної сили генерації АТФ, призводить до ослаблення окислювального фосфорилювання таким чином, що більша частина енергії розсіюється у вигляді тепла і зменшується кількість макроерг. Роз'єднання окислення та фосфорилювання під впливом адреналіну використовується клітинами гомойотермних організмів для підвищення теплопродукції за підтримки постійної температури тіла під час охолодження або її підвищення при лихоманці. Значні зміни у структурі мітохондрій та енергетичному метаболізмі спостерігаються при тиреотоксикозі. Ці зміни спочатку оборотні, але після певної риси стають незворотними: мітохондрії фрагментуються, розпадаються або набухають, втрачають кристи, перетворюючись на вакуолі, і зрештою накопичують такі речовини, як гіалін, феритин, кальцій, ліпофусцин. У хворих на цингу відбувається злиття мітохондрій з утворенням хондріосфер, можливо, внаслідок пошкодження мембран пероксидними сполуками. Значні ушкодження мітохондрій виникають під дією іонізуючого випромінювання, в ході перетворення нормальної клітини на злоякісну.
Мітохондрії є потужним депо іонів кальцію, де його концентрація на кілька порядків перевищує таку в цитоплазмі. При пошкодженні мітохондрій кальцій виходить у цитоплазму, зумовлюючи активізацію протеїназ із пошкодженням внутрішньоклітинних структур та порушенням функцій відповідної клітини, наприклад, кальцієві контрактури або навіть “кальцієву смерть” у нейронах. Внаслідок порушення функціональної здатності мітохондрій різко збільшується утворення вільнорадикальних пероксидних сполук, які мають дуже високу реакційну здатність і тому ушкоджують важливі компоненти клітини – нуклеїнові кислоти, білки та ліпіди. Це спостерігається при так званому оксидаційному стресі і може мати негативні наслідки для існування клітини. Так, пошкодження зовнішньої мембрани мітохондрії супроводжується виходом у цитоплазму речовин, що містяться в міжмембранному просторі, насамперед цитохрому С та деяких інших БАВ, які запускають ланцюгові реакції, що є причиною запрограмованої загибелі клітини – апоптозу. Пошкоджуючи ДНК мітохондрій, вільнорадикальні реакції спотворюють генетичну інформацію, необхідну для утворення деяких ферментів дихального ланцюга, які продукуються саме в мітохондріях. Це призводить до ще більшого порушення окисних процесів. У цілому нині власний генетичний апарат мітохондрій проти генетичним апаратом ядра гірше захищений від шкідливих впливів, здатних змінювати закодовану у ньому генетичну інформацію. Як результат – виникає порушення функцій мітохондрій протягом життя, наприклад, у процесі старіння, при злоякісній трансформації клітини, а також на тлі спадкових мітохондріальних хвороб, пов'язаних із мутацією ДНК мітохондрій у яйцеклітині. В даний час описано понад 50 мітохондріальних мутацій, що викликають спадкові дегенеративні хвороби нервової та м'язової систем. Вони передаються дитині виключно від матері, тому що мітохондрії сперматозоїда не входять до складу зиготи і, відповідно, нового організму.
Порушення збереження та передачі генетичної інформації
Ядро клітини містить більшу частину генетичної інформації, і тим самим забезпечує її нормальне функціонування. За допомогою вибіркової експресії генів воно координує роботу клітини в інтерфазі, зберігає генетичну інформацію, відтворює та передає генетичний матеріал у процесі поділу клітин. У ядрі відбуваються реплікація ДНК та транскрипція РНК. Різні патогенні фактори, такі як ультрафіолетове та іонізуюче випромінювання, вільнорадикальне окиснення, хімічні речовини, віруси, здатні пошкоджувати ДНК. Підраховано, що кожна клітина теплокровної тварини за 1 добу. втрачає понад 10 000 підстав. Сюди слід додати порушення під час копіювання під час поділу. За збереження цих ушкоджень клітина була б здатна вижити. Захист полягає у існуванні потужних систем репарації, таких як ультрафіолетова ендонуклеаза, система репаративної реплікації та рекомбінаційного відновлення, які заміняють порушення ДНК. Генетичні дефекти в репаративних системах викликають розвиток хвороб, зумовлених підвищеною чутливістю до факторів, що ушкоджують ДНК. Це пігментна ксеродерма, а також деякі синдроми прискореного старіння, що супроводжуються підвищеною схильністю до злоякісних пухлин.
Система регуляції процесів реплікації ДНК, транскрипції інформаційною РНК (іРНК), трансляції генетичної інформації з нуклеїнових кислот у структуру білків досить складна та багаторівнева. Крім регуляторних каскадів, що запускають дію факгор транскрипції загальною кількістю понад 3000, які активують певні гени, існує ще багаторівнева регуляторна система, опосередкована малими молекулами РНК (інтерферуючі РНК; РНКи). Геном людини, що складається приблизно з 3 млрд пуринових та піримідинових основ, містить лише 2% структурних генів, які відповідають за синтез білків. Інші забезпечують синтез регуляторних РНК, які одночасно з факторами транскрипції активують або блокують роботу структурних генів на рівні ДНК у хромосомах або впливають на процеси трансляції матричної РНК (мРНК) під час утворення молекули поліпептиду в цитоплазмі. Порушення генетичної інформації може відбуватися на рівні структурних генів, так і регуляторної частини ДНК з відповідними проявами у вигляді різноманітних спадкових захворювань.
Останнім часом велику увагу привертають зміни в генетичному матеріалі, які відбуваються у процесі індивідуального розвитку організму та пов'язані з гальмуванням або активацією певних ділянок ДНК та хромосом внаслідок їх метилювання, ацетилювання та фосфорилювання. Ці зміни зберігаються тривалий час, іноді - протягом усього життя організму від ембріогенезу до старості, і отримали назву епігеномної спадковості.
Розмноженню клітин із зміненою генетичною інформацією перешкоджають також системи (чинники) контролю мітотичного циклу. Вони взаємодіють з циклінзалежними протеїнкіназами та їх каталітичними субодиницями - циклінами - і блокують проходження клітиною повного мітотичного циклу, зупиняючи розподіл на кордоні між пресинтетичною та синтетичною фазами (блок G1/S) аж до завершення репарації ДНК, а у разі її неможливості - клітини. До таких факторів відноситься ген р53, мутація якого спричиняє втрату контролю над проліферацією трансформованих клітин; вона спостерігається майже 50 % випадків раку в людини. Другий контрольний пункт проходження мітотичного циклу знаходиться на межі G2/M. Тут контролюється правильність розподілу хромосомного матеріалу між дочірніми клітинами в мітозі або мейозі за допомогою комплексу механізмів, що контролюють клітинне веретено, центр та центроміри (кінетохори). Неефективність цих механізмів призводить до порушення розподілу хромосом або їх частин, що проявляється відсутністю будь-якої хромосоми в одній з дочірніх клітин (анеуплоїдія), наявністю зайвої хромосоми (поліплоїдія), відривом частини хромосоми (делеція) та перенесенням її на іншу хромосому . Такі процеси дуже часто спостерігаються при розмноженні злоякісно перероджених та трансформованих клітин. Якщо це відбувається під час мейозу зі статевими клітинами, то призводить або до загибелі плода на ранньому етапі ембріонального розвитку, або до народження організму з хромосомною хворобою.
Неконтрольоване розмноження клітин під час зростання пухлин виникає як результат мутації в генах, які контролюють проліферацію клітин та отримали назву онкогенів. Серед більш ніж 70 відомих в даний час онкогенів більша частина відноситься до компонентів регуляції росту клітини, частина представлена факторами транскрипції, що регулюють активність генів, а також факторами, які гальмують поділ і зростання клітин. Ще одним фактором, що обмежує надмірну експансію (поширення) проліферуючих клітин, є скорочення кінців хромосом - теломер, які не здатні в результаті суто стеричного взаємодії повністю реплікуватися, тому після кожного поділу клітини теломери коротшають на певну частину підстав. Таким чином, клітини дорослого організму, що проліферують, після певної кількості поділів (зазвичай від 20 до 100 залежно від виду організму та його віку) вичерпують довжину теломери і подальша реплікація хромосом припиняється. Це явище не виникає у сперматозогенному епітелії, ентероцитах та ембріональних клітинах завдяки наявності ферменту теломерази, що відновлює довжину теломер після кожного поділу. У більшості клітин дорослих організмів теломераза заблокована, але, на жаль, вона активована у клітинах пухлин.
Зв'язок між ядром та цитоплазмою, транспорт речовин в обох напрямках здійснюються через пори в ядерній мембрані за участю спеціальних транспортних систем із споживанням енергії. Таким чином транспортуються до ядра енергетичні та пластичні речовини, сигнальні молекули (чинники транскрипції). Зворотний потік виносить у цитоплазму молекули іРНК та транспортної РНК (тРНК), рибосоми, необхідні синтезу білка в клітині. Цей шлях транспорту речовин притаманний і вірусам, зокрема таким, як ВІЛ. Вони переносять свій генетичний матеріал у ядро клітини-господаря з подальшим включенням його в геном господаря та перенесенням новоутвореної вірусної РНК у цитоплазму для подальшого синтезу білків нових вірусних частинок.
Порушення процесів синтезу
p align="justify"> Процеси синтезу білків відбуваються в цистернах ендоплазматичної мережі, тісно пов'язаних з порами в ядерній мембрані, через які в ендоплазматичну мережу надходять рибосоми, тРНК та іРНК. Тут здійснюється синтез поліпептидних ланцюжків, які надалі набувають свого остаточного вигляду в агранулярній ендоплазматичній мережі та пластинчастому комплексі (комплексі Гольджі), де піддаються трансляційної модифікації та з'єднанню з молекулами вуглеводів і ліпідів. Новоутворені білкові молекули не залишаються на місці синтезу, а за допомогою складного регульованого процесу, що зветься протеїнкінезисуактивно переносяться до тієї ізольованої частини клітини, де вони виконуватимуть призначену їм функцію. При цьому дуже важливим етапом є структурування перенесеної молекули відповідну просторову конфігурацію, здатну виконувати властиву їй функцію. Таке структурування відбувається за допомогою спеціальних ферментів або на матриці спеціалізованих білкових молекул - шаперонів, які допомагають білковій молекулі, новоствореній або зміненій внаслідок зовнішнього впливу, набути правильної тривимірної структури. У разі несприятливого впливу на клітину, коли виникає можливість порушення структури білкових молекул (наприклад, при підвищенні температури тіла, інфекційному процесі, інтоксикації) концентрація шаперонів у клітині різко збільшується. Тому такі молекули отримали ще назву стрес-білків, або білків теплового шоку. Порушення структурування білкової молекули призводить до утворення хімічно інертних конгломератів, які відкладаються в клітині або поза нею при амілоїдозі, хворобі Альцгеймера та ін. будуть дефектними. Ця ситуація виникає при так званих пріонових хворобах (скріпи у овець, сказ корів, куру, хвороба Крейтцфельдта-Якоба у людини), коли дефект одного з мембранних білків нервової клітини зумовлює подальше скупчення інертних мас усередині клітини та порушення її життєдіяльності.
Порушення процесів синтезу в клітині може відбуватися на різних його етапах: транскрипція РНК в ядрі, трансляція поліпептидів у рибосомах, посттрансляційна модифікація, гіперметилювання та глікозилювання бежевої молекули, транспорт та розподіл білків у клітині та виведення їх назовні. При цьому можна спостерігати збільшення або зменшення кількості рибосом, розпад полірибосом, розширення цистерн гранулярної ендоплазматичної мережі, втрату рибосом, утворення везикул і вакуолей. Так, при отруєнні блідою поганкою ушкоджується фермент РНК-полімераза, що порушує транскрипцію. Дифтерійний токсин, інактивуючи фактор елонгації, порушує процеси трансляції, зумовлюючи ушкодження міокарда. Причиною порушення синтезу деяких специфічних білкових молекул можуть бути інфекційні агенти. Наприклад, герпесвіруси гальмують синтез та експресію молекул антигенів ГКГС, що дозволяє їм частково уникнути імунного контролю, бацили чуми – синтез медіаторів гострого запалення. Поява незвичайних білків може призупиняти їхній подальший розпад і призводити до накопичення інертного або навіть токсичного матеріалу. Цьому певною мірою може сприяти порушення процесів розпаду.
Порушення процесів розпаду
Поруч із синтезом білка у клітині безупинно відбувається його розпад. У нормальних умовах це має важливе регуляторне та формотворне значення, наприклад, під час активації неактивних форм ферментів, білкових гормонів, білків мітотичного циклу. Нормальне зростання та розвиток клітини потребують тонко контрольованого балансу між синтезом та деградацією білків та органел. Однак у процесі синтезу білків внаслідок помилок у роботі синтезуючого апарату, аномального структурування білкової молекули, її ушкодження хімічними та бактеріальними агентами постійно утворюється досить багато дефектних молекул. За деякими оцінками, їхня частка становить близько третини всіх синтезованих білків.
Клітини ссавців мають кілька головних шляхів руйнування білків:через лізосомальні протеази (пентидгідролази), кальційзалежні протеїнази (ендопептидази) та систему протеасом. Крім того, є ще й спеціалізовані протеїнази, наприклад, каспази. Основною органелою, де відбувається деградація речовин в еукаріотичних клітинах, є лізосома, що містить численні гідролітичні ферменти. Внаслідок процесів ендоцитозу та різних типів аутофагії в лізосомах та фаголізосомах руйнуються як дефектні білкові молекули, так і цілі органели: пошкоджені мітохондрії, ділянки плазматичної мембрани, деякі екстрацелюлярні білки, вміст секреторних гранул.
Важливим механізмом деградації білків є протеасома - мультиката-літична протеїназна структура складної будови, що локалізується в цитозолі, ядрі, ендоплазматичній мережі та на мембрані клітини. Ця ферментна система відповідає за руйнування пошкоджених білків, а також здорових білків, які мають бути видалені для нормального функціонування клітини. При цьому білки, які підлягають руйнуванню, попередньо з'єднуються зі специфічним поліпептидом убіквітіном. Однак частково в протеасомах можуть руйнуватися і неубіквітовані білки. Розпад білкової молекули в протеасомах до коротких поліпептидів (процесинг) з подальшою їх презентацією спільно з молекулами ГКГС І типу є важливою ланкою у здійсненні імунного контролю антигенного гомеостазу організму. При ослабленні функції протеасом відбувається акумуляція пошкоджених та непотрібних білків, що супроводжує старіння клітини. Порушення деградації циклінзалежних білків призводить до порушення клітинного поділу, деградації секреторних білків – до розвитку цистофіброзу. І навпаки, підвищення функції протеасом супроводжує виснаження організму (СНІД, рак).
При генетично обумовлених порушеннях деградації білків організм нежиттєздатний та гине на ранніх стадіях ембріогенезу. Якщо ж порушується розпад жирів чи вуглеводів, то виникають хвороби накопичення (тезаурісмози). При цьому всередині клітини накопичується надмірна кількість певних речовин або продуктів їхнього неповного розпаду – ліпідів, полісахаридів, що суттєво ушкоджує функцію клітини. Найчастіше це спостерігається в епітеліоцитах печінки (гепатоцитах), нейронах, фібробластах та макрофагоцитах.
Набуті порушення процесів розпаду речовин можуть виникати як результат патологічних процесів (наприклад, білкової, жирової, вуглеводної та пігментної дистрофії) та супроводжуватися утворенням незвичних речовин. Порушення в системі лізосомного протеолізу призводять до зниження адаптації при голодуванні або підвищеному навантаженні, до виникнення деяких ендокринних дисфункцій – зниження рівня інсуліну, тиреоглобуліну, цитокінів та їх рецепторів. Порушення деградації білків уповільнюють швидкість загоєння ран, зумовлюють розвиток атеросклерозу, впливають на імунну відповідь. При гіпоксії, зміні внутрішньоклітинного pH, променевому ураженні, що характеризуються посиленою пероксидацією мембранних ліпідів, а також під впливом лізосомотропних речовин - ендотоксинів бактерій, метаболітів токсичних грибів (спорофузарин), кристалів оксиду кремнію - змінюється стабільність мембрани лізосом що викликає руйнування структур клітини та її загибель.
Запрошуємо Вас ознайомитись з матеріалами та .
: целюлозна оболонка, мембрана, цитоплазма з органоїдами, ядро, вакуолі з клітинним соком.Наявність пластид – головна особливість рослинної клітини.
Функції клітинної оболонки- Визначає форму клітини, захищає від факторів зовнішнього середовища.
Плазматична мембрана- тонка плівка, що складається з взаємодіючих молекул ліпідів та білків, відмежовує внутрішній вміст від зовнішнього середовища, забезпечує транспорт у клітину води, мінеральних та органічних речовин шляхом осмосу та активного перенесення, а також видаляє продукти життєдіяльності.
Цитоплазма- внутрішнє напіврідке середовище клітини, в якому розташоване ядро та органоїди, забезпечує зв'язок між ними, бере участь в основних процесах життєдіяльності.
Ендоплазматична мережа- мережу розгалужених каналів у цитоплазмі. Вона бере участь у синтезі білків, ліпідів та вуглеводів, у транспорті речовин. Рибосоми – тільця, розташовані на ЕПС або в цитоплазмі, складаються з РНК та білка, беруть участь у синтезі білка. ЕПС та рибосоми - єдиний апарат синтезу та транспорту білків.
Мітохондрії– органоїди, відмежовані від цитоплазми двома мембранами. Вони окислюються органічні речовини і синтезуються молекули АТФ з участю ферментів. Збільшення поверхні внутрішньої мембрани, де розташовані ферменти з допомогою христ. АТФ - багата на енергію органічна речовина.
Пластиди(хлоропласти, лейкопласти, хромопласти), їх вміст у клітині – головна особливість рослинного організму. Хлоропласти - пластиди, що містять зелений пігмент хлорофіл, який поглинає енергію світла та використовує її на синтез органічних речовин із вуглекислого газу та води. Відмежування хлоропластів від цитоплазми двома мембранами, численні вирости – грани на внутрішній мембрані, в яких розташовані молекули хлорофілу та ферменти.
Комплекс Гольджі- Система порожнин, відмежованих від цитоплазми мембраною. Нагромадження в них білків, жирів та вуглеводів. Здійснення на мембранах синтезу жирів та вуглеводів.
Лізосоми- тільця, відмежовані від цитоплазми однією мембраною. Ферменти, що містяться в них, прискорюють реакцію розщеплення складних молекул до простих: білків до амінокислот, складних вуглеводів до простих, ліпідів до гліцерину і жирних кислот, а також руйнують відмерлі частини клітини, цілі клітини.
Вакуолі- Порожнини в цитоплазмі, заповнені клітинним соком, місце накопичення запасних поживних речовин, шкідливих речовин; вони регулюють вміст води у клітині.
Ядро- головна частина клітини, покрита зовні двох мембранною, пронизаною порами ядерною оболонкою. Речовини надходять у ядро і віддаляються з нього через пори. Хромосоми – носії спадкової інформації про ознаки організму, основні структури ядра, кожна з яких складається з однієї молекули ДНК у поєднанні з білками. Ядро – місце синтезу ДНК, і-РНК, р-РНК.

Наявність зовнішньої мембрани, цитоплазми з органоїдами, ядра із хромосомами.
Зовнішня, або плазматична мембрана- відмежовує вміст клітини від навколишнього середовища (інших клітин, міжклітинної речовини), складається з молекул ліпідів та білка, забезпечує зв'язок між клітинами, транспорт речовин у клітину (піноцитоз, фагоцитоз) та з клітини.
Цитоплазма- внутрішнє напіврідке середовище клітини, що забезпечує зв'язок між розташованими в ній ядром та органоїдами. У цитоплазмі протікають основні процеси життєдіяльності.
Органоїди клітини:
1) ендоплазматична мережа (ЕПС)- система розгалужених канальців, бере участь у синтезі білків, ліпідів і вуглеводів, у транспорті речовин у клітині;
2) рибосоми- тільця, що містять рРНК, розташовані на ЕПС та в цитоплазмі, беруть участь у синтезі білка. ЕПС та рибосоми - єдиний апарат синтезу та транспорту білка;
3) мітохондрії- "силові станції" клітини, відмежовані від цитоплазми двома мембранами. Внутрішня утворює кристи (складки), що збільшують її поверхню. Ферменти на кристалах прискорюють реакції окислення органічних речовин та синтезу молекул АТФ, багатих на енергію;
4) комплекс Гольджі- група порожнин, відмежованих мембраною від цитоплазми, заповнених білками, жирами та вуглеводами, які або використовуються у процесах життєдіяльності, або видаляються з клітини. На мембранах комплексу здійснюється синтез жирів та вуглеводів;
5) лізосоми- тільця, заповнені ферментами, прискорюють реакції розщеплення білків до амінокислот, ліпідів до гліцерину та жирних -. кислот, полісахаридів до моносахаридів. У лізосомах руйнуються відмерлі частини клітини, цілі та клітини.
Клітинні включення- накопичення запасних поживних речовин: білків, жирів та вуглеводів.
Ядро- Найважливіша частина клітини. Воно покрите двомембранною оболонкою з порами, через які одні речовини проникають у ядро, а інші надходять до цитоплазми. Хромосоми – основні структури ядра, носії спадкової інформації про ознаки організму. Вона передається у процесі поділу материнської клітини дочірнім клітинам, і з статевими клітинами - дочірнім організмам. Ядро – місце синтезу ДНК, іРНК, рРНК.
Завдання:
Поясніть, чому органоїди називають спеціалізованими структурами клітини?
Відповідь:органоїди називають спеціалізованими структурами клітини, оскільки вони виконують суворо певні функції, у ядрі зберігається спадкова інформація, у мітохондріях синтезується АТФ, у хлоропластах протікає фотосинтез тощо.
Якщо у Вас є питання щодо цитології, то Ви можете звернутися за допомогою до
Зв'язок організму з довкіллям, з фізико-хімічної погляду, є відкриту систему, т. е. систему, де біохімічні процеси йдуть постійно. Вихідні речовини надходять із навколишнього середовища, а речовини, що утворюються також безперервно, виносяться зовні. Рівновага між швидкістю і концентрацією продуктів різноспрямованих реакцій в організмі є умовним, уявним, тому що надходження і винесення речовин не припиняються. Безперервний зв'язок із навколишнім середовищем і дозволяє розглядати живий організм як відкриту систему.
Для всіх живих клітин джерелом енергії є Сонце. Рослинні клітини вловлюють енергію сонячного світла з допомогою хлорофілу, використовуючи її реакцій асиміляції у процесі фотосинтезу. Клітини тварин, грибів, бактерій використовують сонячну енергію побічно при розщепленні органічних речовин, синтезованих земною рослиною.
Частина поживних речовин клітини розщеплюється в процесі дихання клітин, поставляючи таким чином енергію, необхідну для різного роду клітинної активності. Протікає цей процес у органелах, званих мітохондріями. Мітохондрія складається з двох мембран: зовнішньої, що відокремлює органеллу від цитоплазми, і внутрішньої, що утворює численні складики. Основним продуктом дихання є АТФ. Він залишає мітохондрії і використовується як джерело енергії для багатьох хімічних реакцій у цитоплазмі та клітинній мембрані. Якщо для здійснення клітинного дихання потрібно кисень, то дихання називають аеробним, якщо ж реакції йдуть за відсутності кисню, то говорять про анаеробне дихання.
Для будь-якого виду роботи, що виконується в клітині, використовується енергія в одній формі - у формі енергії фосфатних зв'язків АТФ. АТФ – легко рухоме з'єднання. Утворення АТФ відбувається на внутрішній мембрані мітохондрій. АТФ синтезується у всіх клітинах при диханні за рахунок енергії окислення вуглеводів, жирів та інших органічних речовин. У клітинах зелених рослин основна кількість АТФ синтезується у хлоропластах за рахунок сонячної енергії. Вони при фотосинтезі утворюється у багато разів більше АТФ, ніж у мітохондріях. Розкладається АТФ з розривом фосфорно-кисневих зв'язків та виділенням енергії. Це відбувається під дією ферменту АТФази у процесі гідролізу АТФ – приєднання води із відщепленням молекули фосфорної кислоти. В результаті АТФ перетворюється на АДФ, а якщо відщеплюються дві молекули фосфорної кислоти, то на АМФ. Реакція відщеплення кожної грам-молекули кислоти супроводжується визволенням 40 кДж. Це дуже великий вихід енергії, тому фосфорно-кисневі зв'язки АТФ прийнято називати макроергітичними (високоенергетичними).
Використання АТФ у реакціях пластичного обміну здійснюється шляхом їх поєднання з гідролізом АТФ. Молекули різних речовин заряджаються енергією шляхом приєднання звільненої гідролізі фосфорної групи від молекули АТФ, тобто шляхом фосфорилювання.
Особливість фосфатних похідних полягає в тому, що вони не можуть залишити клітину, хоча їх розряджені форми вільно проходять через мембрану. Завдяки цьому фосфорильовані молекули залишаються в клітині доти, доки вони не будуть використані у відповідних реакціях.
Зворотний процес перетворення АДФ на АТФ відбувається шляхом приєднання молекули фосфорної кислоти до АДФ з виділенням води та поглинанням великої кількості енергії.
Таким чином АТФ - універсальне та безпосереднє джерело енергії для діяльності клітини. Це створює єдиний клітинний фонд енергії і уможливлює її перерозподіл і транспортування з одних ділянок клітини до інших.
Перенесення фосфатної групи відіграє у хімічних реакціях типу складання макромолекул з мономерів. Наприклад, амінокислоти можуть з'єднуватись у пептиди, лише будучи попередньо профосфорильованими. Механічні процеси скорочення або руху, перенесення розчиненої речовини проти концентрації градієнта та інші процеси пов'язані з витрачанням енергії, запасеної в АТФ.
Процес енергетичного обміну може бути представлений в такий спосіб. Високомолекулярні органічні речовини в цитоплазмі ферментативно, шляхом гідролізу перетворюються на простіші, з яких вони складаються: білки - на амінокислоти, полі- та дисахариди - на моносахариди (+ глюкозу), жири на гліцерин і жирні кислоти. Окисні процеси відсутні, звільняється мало енергії, яка не використовується і переходить у теплову форму. Більшість клітин насамперед витрачають вуглеводи. Полісахариди (крохмаль у рослин та глікоген у тварин) гідралізуються до глюкози. Окислення глюкози відбувається у три фази: гліколіз, окисне декарбоксилювання (цикл Кребса – цикл лимонної кислоти) та окисне фосфорилювання (дихальний ланцюг). Гліколіз, внаслідок якого одна молекула глюкози розщеплюється на дві молекули піровиноградної кислоти з виділенням двох молекул АТФ, протікає у цитоплазмі. За відсутності кисню піровиноградна кислота перетворюється або на етанол (бродіння), або молочну кислоту (анаеробне дихання).
Якщо гліколіз здійснюється у клітинах тварин, шестивуглецева молекула глюкози розпадається на дві молекули молочної кислоти. Процес цей багатоступінчастий. Його послідовно здійснюють 13 ферментів. При спиртовому бродінні з молекули глюкози утворюється дві молекули етанолу та дві молекули СО2.
Гліколіз - фаза, загальна для анаеробного та аеробного дихання, дві інші здійснюються лише в аеробних умовах. Процес безкисневого окислення, у якому виділяється і використовується лише частина енергії метаболітів, для анаеробних організмів є кінцевим. У присутності ж кисню піровиноградна кислота переходить у мітохондрії, де в результаті цілого ряду послідовних реакцій вона повністю окислюється аеробним шляхом до Н2O та СО2 з одночасним фосфорилуванням АДФ до АТФ. При цьому дві молекули АТФ дає гліколіз, дві – цикл Кребса, 34 – дихальний ланцюг. Чистий вихід при повному окисленні однієї молекули глюкози до Н2О та СО2 становить 38 молекул.
Таким чином, у аеробних організмів остаточний розпад органічних речовин здійснюється шляхом окислення їх киснем повітря до простих неорганічних: СО2 та Н2О. Процес цей протікає на кристалах мітохондрій. При цьому виділяється максимальна кількість вільної енергії, значна частина якої резервується молекулами АТФ. Легко бачити, що аеробне окислення найбільше забезпечує клітину вільної енергією.
В результаті катаболізму в клітині накопичуються багаті на енергію молекули АТФ, а в зовнішнє середовище виділяється СО2 і надмірна кількість води.
Молекули цукрів, які не потрібні для дихання, можуть запасатися в клітці. Надлишкові ліпіди або розщеплюються, після чого продукти їх розщеплення надходять у мітохондрії як субстрат для дихання, або відкладаються про запас у цитоплазмі у вигляді жирових крапель. З амінокислот, що надходять у клітину, будуються білки. Синтез білка відбувається в органелах, які називаються рибосомами. Кожна рибосома складається з двох субчастинок - великої та малої: до складу обох субчасток входять білкові молекули та молекули РНК.
Рибосоми часто бувають прикріплені до особливої системи мембран, що складається з цистерн та бульбашок, - до так званого ендоплазматичного ретикулуму (ЕР); в клітинах, що виробляють багато білка, ендоплазматичний ретикулум часто розвинений дуже добре і весь усіяний рибосомами. Деякі ферменти ефективні лише тому випадку, якщо вони прикріплені до мембрани. Тут перебуває більшість ферментів, що у синтезі ліпідів. Таким чином, ендоплазматичний ретикулум - це своєрідний робочий стіл клітини.
Крім того, ЕР ділить цитоплазму на окремі відсіки, або компартменти, тобто роз'єднує різні хімічні процеси, що одночасно протікають у цитоплазмі, і тим самим знижує ймовірність того, що ці процеси будуть заважати один одному.
Продукти, що часто утворюються даною клітиною, використовуються поза цією клітиною. У подібних випадках синтезовані на рибосомах білки проходять через мембрани ендоплазматичного ретикулуму і упаковуються в мембранні бульбашки, що формуються навколо них, які потім відшнуровуються від ЕР. Ці бульбашки, уплощаясь і укладаючись один на одного, як млинці в стопці, утворюють характерну структуру, яка називається комплексом Гольджі, або апаратом Гольджі. Під час свого перебування в апараті Гольджі білки зазнають певних змін. Коли їм настає час залишити клітину, мембранні бульбашки зливаються з клітинної мембраною і спорожняються, виливаючи свій вміст назовні, т. е. секреція відбувається шляхом екзоцитозу.
В апараті Гольджі утворюються також лізосоми – мембранні мішечки, що містять травні ферменти. З'ясування того, як клітина виробляє, упаковує та експортує деякі білки, а також як вона «дізнається», які білки їй слід зберігати для себе, становить один із найцікавіших розділів сучасної цитології.
Мембрани будь-якої клітини безперервно переміщуються і видозмінюються. Мембрани ЕР повільно переміщаються клітиною. Окремі ділянки цих мембран відокремлюються і утворюють бульбашки, які на якийсь час стають частиною апарату Гольджі, а потім, в процесі екзоцитозу, зливаються з клітинною мембраною.
Пізніше мембранний матеріал повертається до цитоплазми, де він використовується знову.
Теорія до завдання 5 з ЄДІ з біології
Будова клітини. Взаємозв'язок будови та функцій частин та органоїдів клітини – основа її цілісності
Будова клітини
Будова прокаріотичних та еукаріотичних клітин
Основними структурними компонентами клітин є плазматична мембрана, цитоплазма та спадковий апарат. Залежно від особливостей організації розрізняють два основні типи клітин: прокаріотичні та еукаріотичні. Головною відмінністю прокаріотів від еукаріотичних клітин є організація їх спадкового апарату: у прокаріотів він знаходиться безпосередньо в цитоплазмі (ця область цитоплазми називається нуклеоїдом) і не відокремлений від неї мембранними структурами, тоді як у еукаріотів більша частина ДНК зосереджена в ядрі, оточеному подвійною мембраною. Крім того, генетична інформація прокаріотів, що знаходиться в нуклеоїді, записана в кільцевій молекулі ДНК, а у еукаріотів молекули ДНК незамкнуті.
На відміну від еукаріотів, цитоплазма прокаріотичних клітин містить також невелику кількість органоїдів, тоді як для еукаріотів характерна значна різноманітність цих структур.
Будова та функції біологічних мембран
Будова біомембрани.Мембрани, що обмежують клітини та мембранні органоїди еукаріотичних клітин, мають загальний хімічний склад та будову. До їх складу входять ліпіди, білки та вуглеводи. Ліпіди мембрани представлені в основному фосфоліпідами та холестерином. Більшість білків мембран відноситься до складних білків, наприклад, глікопротеїнів. Вуглеводи не зустрічаються в мембрані самостійно, вони пов'язані з білками та ліпідами. Товщина мембран складає 7-10 нм.
Відповідно до загальноприйнятої в даний час рідинно-мозаїчної моделі будови мембран, ліпіди утворюють подвійний шар, або ліпідний бішар, В якому гідрофільні «головки» молекул ліпідів звернені назовні, а гідрофобні «хвости» заховані всередину мембрани. Ці «хвости» завдяки своїй гідрофобності забезпечують поділ водних фаз внутрішнього середовища клітини та її оточення. З ліпідами з допомогою різних типів взаємодії пов'язані білки. Частина білків розташована на поверхні мембрани. Такі білки називають периферичними, або поверхневими. Інші білки частково або повністю занурені в мембрану – це інтегральні,або занурені білки. Білки мембран виконують структурну, транспортну, каталітичну, рецепторну та інші функції.
Мембрани не схожі на кристали, їх компоненти постійно перебувають у русі, внаслідок чого між молекулами ліпідів виникають розриви - пори, через які в клітину можуть потрапляти або залишати різні речовини.
Біологічні мембрани розрізняються за розташуванням у клітині, хімічному складі та виконуваних функцій. Основні типи мембран - плазматична та внутрішні. Плазматична мембранамістить близько 45% ліпідів (у т. ч. гліколіпідів), 50% білків та 5% вуглеводів. Ланцюжки вуглеводів, що входять до складу складних білків-глікопротеїнів та складних ліпідів-гліколіпідів, виступають над поверхнею мембрани. Глікопротеїни плазмалеми надзвичайно специфічні. Так, наприклад, по них відбувається взаємне впізнавання клітин, у тому числі сперматозоїда та яйцеклітини.
На поверхні тварин клітин вуглеводні ланцюжки утворюють тонкий поверхневий шар. глікоколікс.Він виявлений майже у всіх тваринних клітинах, але ступінь його вираженості неоднакова (10-50 мкм). Глікокалікс забезпечує безпосередній зв'язок клітини із зовнішнім середовищем, у ньому відбувається позаклітинне травлення; у глікокаліксі розміщені рецептори. Клітини бактерій, рослин та грибів, крім плазмалеми, оточені ще й клітинними оболонками.
Внутрішні мембраниеукаріотичних клітин розмежовують різні частини клітини, утворюючи своєрідні «відсіки» компартментищо сприяє поділу різних процесів обміну речовин та енергії. Вони можуть відрізнятися за хімічним складом і функцій, що виконуються, але загальний план будівлі у них зберігається.
Функції мембран:
- Обмежує.Полягає в тому, що вони відокремлюють внутрішній простір клітини від зовнішнього середовища. Мембрана є напівпроникною, тобто її вільно долають ті речовини, які необхідні клітині, при цьому існують механізми транспорту необхідних речовин.
- Рецепторна.Пов'язана насамперед із сприйняттям сигналів навколишнього середовища та передачею цієї інформації всередину клітини. За цю функцію відповідають спеціальні білки-рецептори. Мембранні білки відповідають ще й за клітинне впізнавання за принципом "свій-чужий", а також за утворення міжклітинних сполук, найбільш вивченими з яких є синапси нервових клітин.
- Каталітична.На мембранах розташовані численні ферментні комплекси, унаслідок чого ними відбуваються інтенсивні синтетичні процеси.
- Енерготрансформуюча.Пов'язана з утворенням енергії, її запасанням у вигляді АТФ та витрачанням.
- Компартменталізація.Мембрани розмежовують також простір усередині клітини, поділяючи цим вихідні речовини реакції і ферменти, які можуть здійснювати відповідні реакції.
- Утворення міжклітинних контактів.Незважаючи на те, що товщина мембрани настільки мала, що її неможливо розрізнити неозброєним оком, вона, з одного боку, є досить надійним бар'єром для іонів і молекул, особливо водорозчинних, а з іншого забезпечує їх перенесення в клітину і назовні.
- Транспортні.

Мембранний транспорт.У зв'язку з тим, що клітини як елементарні біологічні системи є відкритими системами для забезпечення обміну речовин і енергії, підтримки гомеостазу, росту, подразливості та інших процесів потрібне перенесення речовин через мембрану — мембранний транспорт. В даний час транспорт речовин через мембрану клітини ділять на активний, пасивний, ендо- та екзоцитоз.
Пасивний транспорт- Це вид транспорту, який відбувається без витрати енергії від більшої концентрації до меншої. Розчинні в ліпідах невеликі неполярні молекули (О2, СО2) легко проникають у клітину шляхом простий дифузії. Нерозчинні ж у ліпідах, у тому числі заряджені невеликі частинки, підхоплюються білками-переносниками або проходять через спеціальні канали (глюкоза, амінокислоти, К+, PO 4 3-). Такий вид пасивного транспорту називається полегшеною дифузією. Вода надходить у клітину через пори в ліпідній фазі, а також спеціальними каналами, вистеленими білками. Транспорт води через мембрану називається осмосом.

Осмос має надзвичайно важливе значення в житті клітини, оскільки якщо її помістити в розчин з більш високою концентрацією солей, ніж у клітинному розчині, вода почне виходити з клітини, і об'єм живого вмісту почне зменшуватися. У тварин клітин відбувається з'їджування клітини загалом, а рослинних — відставання цитоплазми від клітинної стінки, що називається плазмолізом. При поміщенні клітини менш концентрований, ніж цитоплазма, розчин, транспорт води відбувається у зворотному напрямку — в клітину. Однак існують межі розтяжності цитоплазматичної мембрани, і тваринна клітина зрештою розривається, а у рослинної цього не дозволяє зробити міцна клітинна стінка. Явище заповнення клітинним вмістом всього внутрішнього простору клітини називається деплазмолізом. Внутрішньоклітинну концентрацію солей слід враховувати при приготуванні лікарських препаратів, особливо для внутрішньовенного введення, оскільки це може призводити до пошкодження клітин крові (для цього використовують фізіологічний розчин із концентрацією 0,9 % хлориду натрію). Це не менш важливо при культивуванні клітин та тканин, а також органів тварин та рослин.

Активний транспортпротікає із витратою енергії АТФ від меншої концентрації речовини до більшої. Він здійснюється за допомогою спеціальних білків-насосів. Білки перекачують через мембрану іони К+, Na+, Са2+ та інші, що сприяє транспорту найважливіших органічних речовин, а також виникненню нервових імпульсів тощо.
Ендоцитоз- це активний процес поглинання речовин клітиною, при якому мембрана утворює вп'ячування, а потім формує мембранні бульбашки. фагосоми, в яких укладені об'єкти, що поглинаються. Потім з фагосомою зливається первинна лізосома і утворюється вторинна лізосома, або фаголізосома, або травна вакуоля. Вміст бульбашки розщеплюється ферментами лізосом, а продукти розщеплення поглинаються та засвоюються клітиною. Неперетравлені залишки видаляються із клітини шляхом екзоцитозу. Розрізняють два основні види ендоцитозу: фагоцитоз та піноцитоз.
Фагоцитоз- Це процес захоплення клітинною поверхнею і поглинання клітиною твердих частинок, а піноцитоз- Рідини. Фагоцитоз протікає переважно у тварин клітинах (одноклітинні тварини, лейкоцити людини), він забезпечує їх харчування, а то й захист організму. Шляхом піноцитозу відбувається поглинання білків, комплексів антиген-антитіла в процесі імунних реакцій і т. д. Однак шляхом піноцитозу або фагоцитозу в клітину також потрапляють багато вірусів. У клітинах рослин та грибів фагоцитоз практично неможливий, оскільки вони оточені міцними клітинними оболонками.

Екзоцитоз- Процес, зворотний ендоцитозу. Таким чином виділяються неперетравлені залишки їжі з травних вакуолей, виводяться необхідні для життєдіяльності клітини та організму загалом речовини. Наприклад, передача нервових імпульсів відбувається завдяки виділенню нейроном, що посилає імпульс, хімічних посередників. медіаторів, а рослинних клітинах так виділяються допоміжні вуглеводи клітинної оболонки.
Клітинні оболонки клітин рослин, грибів та бактерій.Зовні від мембрани клітина може виділяти міцний каркас. клітинну оболонку,або клітинну стінку.
У рослин основу клітинної оболонки становить целюлоза, Упакована в пучки по 50-100 молекул. Проміжки між ними заповнюють вода та інші вуглеводи. Оболонка рослинної клітини пронизана канальцями плазмодесмамичерез які проходять мембрани ендоплазматичної мережі. По плазмодесмам здійснюється транспорт речовин між клітинами. Однак транспорт речовин, наприклад води, може відбуватися і по самих клітинних стінках. Згодом у клітинній оболонці рослин накопичуються різні речовини, у тому числі дубильні або жироподібні, що призводить до здерев'язання або випробування самої клітинної стінки, витіснення води та відмирання клітинного вмісту. Між клітинними стінками сусідніх клітин рослин розташовуються желеподібні прокладки - серединні пластинки, які скріплюють їх між собою та цементують тіло рослини загалом. Вони руйнуються лише в процесі дозрівання плодів та при опаданні листя.

Клітинні стінки клітин грибів утворені хітіном- Вуглеводом, що містить азот. Вони досить міцні і є зовнішнім скелетом клітини, але все ж таки, як і у рослин, перешкоджають фагоцитозу.
У бактерій до складу клітинної стінки входить вуглевод із фрагментами пептидів. муреїн, проте його зміст суттєво відрізняється у різних груп бактерій. Поверх від клітинної стінки можуть виділятися інші полісахариди, що утворюють слизову капсулу, що захищає бактерії від зовнішніх впливів.
Оболонка визначає форму клітини, служить механічною опорою, виконує захисну функцію, забезпечує осмотичні властивості клітини, обмежуючи розтяг живого вмісту і запобігаючи розриву клітини, що збільшується внаслідок надходження води. Крім того, клітинну стінку долають вода і розчинені в ній речовини, перш ніж потрапити до цитоплазми або, навпаки, при виході з неї, при цьому по клітинних стінках транспортується вода швидше, ніж по цитоплазмі.
Цитоплазма
Цитоплазма- Це внутрішній вміст клітини. У неї занурені всі органоїди клітини, ядро та різноманітні продукти життєдіяльності.
Цитоплазма пов'язує всі частини клітини між собою, у ній протікають численні реакції обміну речовин. Цитоплазма відокремлюється від навколишнього середовища та ділиться на відсіки мембранами, тобто клітинам властива мембранна будова. Вона може перебувати у двох станах — золю та гелю. Золь- Це напіврідкий, киселеподібний стан цитоплазми, при якому процеси життєдіяльності протікають найбільш інтенсивно, а гель- Більш щільний, студнеподібний стан, що ускладнює перебіг хімічних реакцій та транспорт речовин.
Рідка частина цитоплазми без органоїдів називається гіалоплазмою. Гіалоплазма, або цитозоль, є колоїдним розчином, в якому знаходиться своєрідна суспензія досить великих частинок, наприклад білків, оточених диполями молекул води. Осадження цієї суспензії немає внаслідок те, що вони мають однаковий заряд і відштовхуються друг від друга.
Органоїди
Органоїдице постійні компоненти клітини, що виконують певні функції.
Залежно від особливостей будови їх поділяють на мембранні та немембранні. Мембранніорганоїди, у свою чергу, відносять до одномембранних (ендоплазматична мережа, комплекс Гольджі та лізосоми) або двомембранних (мітохондрії, пластиди та ядро). Немембраннимиорганоїдами є рибосоми, мікротрубочки, мікрофіламенти та клітинний центр. Прокаріотам із перелічених органоїдів притаманні лише рибосоми.
Будова та функції ядра. Ядро- Великий двомембранний органоїд, що лежить у центрі клітини або на її периферії. Розміри ядра можуть коливатися не більше 3-35 мкм. Форма ядра частіше сферична або еліпсоїдна, проте є також паличкоподібні, веретеноподібні, бобоподібні, лопатеві і навіть сегментовані ядра. Деякі дослідники вважають, що форма ядра відповідає формі самої клітини.
Більшість клітин має одне ядро, але, наприклад, у клітинах печінки і серця їх може бути два, а в ряді нейронів — до 15. Волокна скелетних м'язів містять зазвичай багато ядер, проте вони не є клітинами в повному розумінні цього слова, оскільки утворюються внаслідок злиття кількох клітин.
Ядро оточене ядерною оболонкою, а його внутрішній простір заповнено ядерним соком, або нуклеоплазмою (каріоплазмою), в яку занурені хроматині ядерце. Ядро виконує такі найважливіші функції, як зберігання та передача спадкової інформації, а також контроль життєдіяльності клітини.

Роль ядра у передачі спадкової інформації була переконливо доведена в експериментах із зеленою водоростями ацетабулярією. В єдиній гігантській клітині, що досягає в довжину 5 см, розрізняють капелюшок, ніжку та різозій. При цьому вона містить лише одне ядро, розташоване у ризоїді. У 1930-ті роки І. Хеммерлінг пересадив ядро одного виду ацетабулярії з зеленим забарвленням в різоїд іншого виду, з коричневим забарвленням, у якого було видалено ядро. Через деякий час у рослини з пересадженим ядром виріс новий капелюшок, як у водорості-донора ядра. У той самий час відокремлені від ризоида капелюшок чи ніжка, які містять ядра, через деякий час гинули.

Ядерна оболонкаутворена двома мембранами - зовнішньою та внутрішньою, між якими є простір. Міжмембранний простір повідомляється з порожниною шорсткої ендоплазматичної мережі, а зовнішня мембрана ядра може нести рибосоми. Ядерна оболонка пронизана численними порами, окантованими спеціальними білками. Через пори відбувається транспорт речовин: у ядро потрапляють необхідні білки (в т. ч. ферменти), іони, нуклеотиди та інші речовини, і залишають його молекули РНК, відпрацьовані білки, суб'одиниці рибосом. Таким чином, функціями ядерної оболонки є відокремлення вмісту ядра від цитоплазми, а також регуляція обміну речовин між ядром і цитоплазмою.
Нуклеоплазмоюназивають вміст ядра, в яке занурені хроматин та ядерце. Вона являє собою колоїдний розчин, що за хімічним складом нагадує цитоплазму. Ферменти нуклеоплазми каталізують обмін амінокислот, нуклеотидів, білків та ін. Нуклеоплазма пов'язана з гіалоплазмою через ядерні пори. Функції нуклеоплазми, як і гіалоплазми, полягають у забезпеченні взаємозв'язку всіх структурних компонентів ядра та здійсненні низки ферментних реакцій.
Хроматиномназивають сукупність тонких ниток та гранул, занурених у нуклеоплазму. Виявити його можна лише при фарбуванні, оскільки коефіцієнти заломлення хроматину та нуклеоплазми приблизно однакові. Нитчастий компонент хроматину називають еухроматином, а гранулярний - гетерохроматином. Еухроматин слабо ущільнений, оскільки з нього зчитується спадкова інформація, тоді як спіралізованіший гетерохроматин є генетично неактивним.
Хроматин є структурною видозміною хромосом в ядрі, що не ділиться. Таким чином, хромосоми постійно присутні в ядрі, змінюється лише їх стан залежно від функції, яку виконує ядро в даний момент.
До складу хроматину в основному входять білки-нуклеопротеїни (дезоксирибонуклеопротеїни та рибонуклеопротеїни), а також ферменти, найважливіші з яких пов'язані із синтезом нуклеїнових кислот, та деякі інші речовини.
Функції хроматину складаються, по-перше, у синтезі специфічних для даного організму нуклеїнових кислот, які спрямовують синтез специфічних білків, по-друге, у передачі спадкових властивостей від материнської клітини дочірнім, для чого хроматинові нитки у процесі розподілу упаковуються у хромосоми.
Ядрішко- сферичне, добре помітне під мікроскопом тільце діаметром 1-3 мкм. Воно формується на ділянках хроматину, в яких закодована інформація про структуру рРНК та білки рибосом. Ядро в ядрі часто одне, проте в тих клітинах, де відбуваються інтенсивні процеси життєдіяльності, ядер може бути два і більше. Функції ядерців - синтез рРНК і складання субодиниць рибосом шляхом поєднання рРНК з білками, що надходять з цитоплазми.
Мітохондрії- Двомембранні органоїди округлої, овальної або паличкоподібної форми, хоча зустрічаються і спіралеподібні (у сперматозоїдах). Діаметр мітохондрій становить до 1 мкм, а довжина – до 7 мкм. Простір усередині мітохондрій заповнений матриксом. Матрікс- Це основна речовина мітохондрій. У нього занурені кільцева молекула ДНК та рибосоми. Зовнішня мембрана мітохондрій гладка, вона непроникна багатьом речовин. Внутрішня мембрана має вирости. кристи, що збільшують площу поверхні мембран для протікання хімічних реакцій На поверхні мембрани розташовані численні білкові комплекси, що становлять так званий дихальний ланцюг, а також грибоподібні ферменти АТФ-синтетази. У мітохондріях протікає аеробний етап дихання, під час якого відбувається синтез АТФ.

Пластиди— великі двомембранні органоїди, характерні лише рослинних клітин. Внутрішній простір пластид заповнено крутий, або матриксом. У стромі знаходиться більш менш розвинена система мембранних бульбашок. тилакоїдів, які зібрані в стопки грани, а також власна кільцева молекула ДНК та рибосоми. Розрізняють чотири основні типи пластид: хлоропласти, хромопласти, лейкопласти та пропластиди.
Хлоропласти- Це зелені пластиди діаметром 3-10 мкм, добре помітні під мікроскопом. Вони містяться тільки в зелених частинах рослин – листі, молодих стеблах, квітках та плодах. Хлоропласти в основному мають овальну або еліпсоїдну форми, але можуть бути також чашоподібними, спіралеподібними і навіть лопатевими. Кількість хлоропластів у клітині становить від 10 до 100 штук. Однак, наприклад, у деяких водоростей він може бути один, мати значні розміри та складну форму - тоді його називають хроматофором. В інших випадках кількість хлоропластів може досягати кількох сотень, причому їх розміри невеликі. Забарвлення хлоропластів зумовлене основним пігментом фотосинтезу. хлорофілом, хоча в них містяться і додаткові пігменти. каротиноїди. Каротиноїди стають помітними лише восени, коли хлорофіл у старілому листі руйнується. Основною функцією хлоропластів є фотосинтез. Світлові реакції фотосинтезу протікають на мембранах тилакоїдів, на яких закріплені молекули хлорофілу, а темнові реакції - у стромі, де містяться численні ферменти.

Хромопласти- це жовті, помаранчеві та червоні пластиди, що містять пігменти каротиноїди. Форма хромопластів може також істотно варіювати: вони бувають трубчастими, сферичними, кристалічними та ін.
Лейкопласти- це білі або безбарвні пластиди в основному округлої або овальної форми. Вони поширені в нефотосинтезирующих частинах рослин, наприклад у шкірці листа, бульбах картоплі і т. д. У них відкладаються в запас поживні речовини, найчастіше крохмаль, але в деяких рослин це можуть бути білки або олія.
Пластиди утворюються в рослинних клітинах з пропластид, які є вже в клітинах освітньої тканини і є невеликими двомембранними тільцями. На ранніх етапах розвитку різні види пластид здатні перетворюватися один на одного: при попаданні на світ лейкопласти бульби картоплі та хромопласти коренеплоду моркви зеленіють.
Пластиди та мітохондрії називають напівавтономними органоїдами клітини, оскільки вони мають власні молекули ДНК та рибосоми, здійснюють синтез білка та діляться незалежно від поділу клітин. Ці особливості пояснюються походженням одноклітинних прокаріотичних організмів. Однак «самостійність» мітохондрій і пластид є обмеженою, оскільки їх ДНК містить дуже мало генів для вільного існування, решта інформації закодована в хромосомах ядра, що дозволяє йому контролювати дані органоїди.
Ендоплазматична мережа (ЕПС), або ендоплазматичний ретикулум (ЕР), - це одномембранний органоїд, що є мережею мембранних порожнин і канальців, що займають до 30% вмісту цитоплазми. Діаметр канальців ЕПС становить близько 25-30 нм. Розрізняють два види ЕПС - шорстку і гладку. Шорстка ЕПСнесе рибосоми, у ньому відбувається синтез білків. Гладка ЕПСпозбавлена рибосом. Її функція – синтез ліпідів та вуглеводів, а також транспорт, запасання та знешкодження токсичних речовин. Вона особливо розвинена у тих клітинах, де відбуваються інтенсивні процеси обміну речовин, наприклад у клітинах печінки – гепатоцитах – та волокнах скелетних м'язів. Речовини, синтезовані в ЕПС, транспортуються в апарат Гольджі. В ЕПС відбувається також складання мембран клітини, проте їхнє формування завершується в апараті Гольджі.
Апарат Гольджі,або комплекс Гольджі, - Одномембранний органоїд, утворений системою плоских цистерн, канальців і бульбашок, що відшнуровуються від них. Структурною одиницею апарату Гольджі є диктіосома- стопка цистерн, на один полюс якої приходять речовини з ЕПС, а з протилежного полюса, зазнавши певних перетворень, вони упаковуються в бульбашки і прямують в інші частини клітини. Діаметр цистерн – близько 2 мкм, а дрібних бульбашок – близько 20-30 мкм. Основні функції комплексу Гольджі - синтез деяких речовин та модифікація (зміна) білків, ліпідів та вуглеводів, що надходять з ЕПС, остаточне формування мембран, а також транспорт речовин по клітині, оновлення її структур та утворення лізосом. Свою назву апарат Гольджі отримав на честь італійського вченого Камілло Гольджі, який вперше виявив цей органоїд (1898).

Лізосоми- невеликі одномембранні органоїди до 1 мкм у діаметрі, у яких містяться гідролітичні ферменти, що беруть участь у внутрішньоклітинному травленні. Мембрани лізосом слабопроникні для цих ферментів, тому виконання лізосом своїх функцій відбувається дуже точно і адресно. Так, вони беруть активну участь у процесі фагоцитозу, утворюючи травні вакуолі, а у разі голодування чи пошкодження певних частин клітини перетравлюють їх, не торкаючись інших. Нещодавно було відкрито роль лізосом у процесах клітинної загибелі.
Вакуоль- це порожнина в цитоплазмі рослинних і тваринних клітин, обмежена мембраною і заповнена рідиною. У клітинах найпростіших виявляються травні та скорочувальні вакуолі. Перші беруть участь у процесі фагоцитозу, оскільки у них відбувається розщеплення поживних речовин. Другі забезпечують підтримку водно-сольового балансу з допомогою осморегуляції. У багатоклітинних тварин переважно зустрічаються травні вакуолі.
У рослинних клітинах вакуолі присутні завжди, вони оточені спеціальною мембраною та заповнені клітинним соком. Мембрана, що оточує вакуоль, за хімічним складом, будовою та виконуваними функціями близька до плазматичної мембрани. Клітинний сікявляє собою водний розчин різних неорганічних і органічних речовин, у тому числі мінеральних солей, органічних кислот, вуглеводів, білків, глікозидів, алкалоїдів та ін. Ця частина клітини виконує запасну, видільну, осмотичну, захисну, лізосомну та інші функції, оскільки в ній накопичуються поживні речовини та відходи життєдіяльності, вона забезпечує надходження води та підтримання форми та об'єму клітини, а також містить ферменти розщеплення багатьох компонентів клітини. До того ж біологічно активні речовини вакуолей здатні перешкоджати поїданню цих рослин багатьма тваринами. У ряду рослин за рахунок набухання вакуолей відбувається зростання клітини розтягуванням.
Вакуолі є також і в клітинах деяких грибів і бактерій, проте у грибів вони виконують лише функцію осморегуляції, а у ціанобактерій підтримують плавучість і беруть участь у процесах засвоєння азоту з повітря.
Рибосоми- Невеликі немембранні органоїди діаметром 15-20 мкм, що складаються з двох субодиниць - великий і малий. Субодиниці рибосом еукаріотів збираються в ядерці, а потім транспортуються в цитоплазму. Рибосоми прокаріотів, мітохондрій і пластид менше за величиною, ніж рибосоми еукаріотів. До складу субодиниць рибосом входять рРНК та білки.

Кількість рибосом у клітині може сягати кількох десятків мільйонів: у цитоплазмі, мітохондріях і пластидах вони у вільному стані, але в шорсткої ЕПС — у зв'язаному. Вони беруть участь у синтезі білка, зокрема, здійснюють процес трансляції – біосинтезу поліпептидного ланцюга на молекулі іРНК. На вільних рибосомах синтезуються білки гіалоплазми, мітохондрій, пластид і власні білки рибосом, тоді як на прикріплених до шорсткої ЕПС рибосом здійснюється трансляція білків для виведення з клітин, складання мембран, утворення лізосом і вакуолей.
Рибосоми можуть знаходитися в гіалоплазмі поодинці або збиратися в групи при одночасному синтезі на одній іРНК відразу кількох поліпептидних ланцюгів. Такі групи рибосом називаються полірибосомами, або полісомами.

Мікротрубочки- Це циліндричні порожнисті немембранні органоїди, які пронизують всю цитоплазму клітини. Їх діаметр становить близько 25 нм, товщина стінки – 6-8 нм. Вони утворені численними молекулами білка тубуліна,які спочатку формують 13 ниток, що нагадують намисто, а потім збираються в мікротрубочку. Мікротрубочки утворюють цитоплазматичну мережу, яка надає клітині форму та об'єм, пов'язують плазматичну мембрану з іншими частинами клітини, забезпечують транспорт речовин по клітині, беруть участь у русі клітини та внутрішньоклітинних компонентів, а також у розподілі генетичного матеріалу. Вони входять до складу клітинного центру та органоїдів руху — джгутиків та вій.
Мікрофіламенти,або мікронитки, також є немембранними органоїдами, проте вони мають ниткоподібну форму та утворені не тубуліном, а актином. Вони беруть участь у процесах мембранного транспорту, міжклітинному впізнанні, розподілі цитоплазми клітини та у її русі. У м'язових клітинах взаємодія актинових мікрофіламентів із міозиновими нитками забезпечує скорочення.
Мікротрубочки та мікрофіламенти утворюють внутрішній скелет клітини. цитоскелет. Він є складною мережею волокон, які забезпечують механічну опору для плазматичної мембрани, визначає форму клітини, розташування клітинних органоїдів та його переміщення у процесі поділу клітини.

Клітинний центр- Немембранний органоїд, що знаходиться в тваринних клітинах поблизу ядра; у рослинних клітинах він відсутній. Його довжина становить близько 0.2-0.3 мкм, а діаметр – 0.1-0.15 мкм. Клітинний центр утворений двома центріолями, що лежать у взаємно перпендикулярних площинах, та променистою сфероюз мікротрубочок. Кожна центріоль утворена дев'ятьма групами мікротрубочок, зібраних по три, тобто триплетами. Клітинний центр бере участь у процесах складання мікротрубочок, розподілі спадкового матеріалу клітини, а також в утворенні джгутиків та вій.

Організації руху. Джгутикиі віїявляють собою вирости клітини, вкриті плазмалемою. Основу цих органоїдів становлять дев'ять пар мікротрубочок, розташованих по периферії, і дві вільні мікротрубочки в центрі. Мікротрубочки пов'язані між собою різними білками, що забезпечують їхнє узгоджене відхилення від осі - коливання. Коливання енергозалежні, тобто цей процес витрачається енергія макроергічних зв'язків АТФ. Відновлення втрачених джгутиків та вій є функцією базальних тілець, або кінетосом, розташованих у їх основі.
Довжина вій становить близько 10-15 нм, а джгутиків - 20-50 мкм. За рахунок строго спрямованих рухів джгутиків і вій здійснюється не тільки рух одноклітинних тварин, сперматозоїдів та ін, але й відбувається очищення дихальних шляхів, просування яйцеклітини по маткових трубах, оскільки всі ці частини організму людини вистелені війчастим епітелієм.

Увімкнення
Увімкнення- це непостійні компоненти клітини, які утворюються та зникають у процесі її життєдіяльності. До них відносять як запасні речовини, наприклад, зерна крохмалю або білка в рослинних клітинах, гранули глікогену в клітинах тварин і грибів, волютину у бактерій, краплі жиру у всіх типах клітин, так і відходи життєдіяльності, зокрема неперетравлені в результаті фагоцитозу залишки їжі , що утворюють так звані залишкові тільця

Взаємозв'язок будови та функцій частин та органоїдів клітини – основа її цілісності
Кожна частина клітини, з одного боку, є відокремленою структурою зі специфічним будовою і функціями, з другого — компонентом складнішої системи, званої клітиною. Більша частина спадкової інформації еукаріотичної клітини зосереджена в ядрі, проте саме ядро не в змозі забезпечити її реалізацію, оскільки для цього необхідні як мінімум цитоплазма, яка виступає як основна речовина, і рибосоми, на яких відбувається цей синтез. Більшість рибосом розташовані на гранулярній ендоплазматичній мережі, звідки білки найчастіше транспортуються в комплекс Гольджі, а потім після модифікації — у ті частини клітини, для яких вони призначені, або виводяться назовні. Мембранні упаковки білків та вуглеводів можуть вбудовуватись у мембрани органоїдів та цитоплазматичну мембрану, забезпечуючи їх постійне оновлення. Від комплексу Гольджі відшнуровуються також виконують найважливіші функції лізосоми та вакуолі. Наприклад, без лізосом клітини швидко перетворилися б на своє образне звалище відпрацьованих молекул і структур.
Протікання всіх цих процесів вимагає енергії, що виробляється мітохондріями, а рослин — і хлоропластами. І хоча ці органоїди є відносно автономними, тому що мають власні молекули ДНК, частина їх білків однаково кодується ядерним геномом і синтезується у цитоплазмі.
Таким чином, клітина є нерозривною єдністю складових її компонентів, кожен з яких виконує свою унікальну функцію.
Обмін речовин та перетворення енергії – властивості живих організмів. Енергетичний та пластичний обмін, їх взаємозв'язок. Стадії енергетичного обміну. Бродіння та дихання. Фотосинтез, значення, космічна роль. Фази фотосинтезу. Світлові та темнові реакції фотосинтезу, їх взаємозв'язок. Хемосинтез. Роль хемосинтезуючих бактерій на Землі
Обмін речовин та перетворення енергії - властивості живих організмів
Клітину можна уподібнити мініатюрній хімічній фабриці, де відбуваються сотні і тисячі хімічних реакцій.
Обмін речовин- Сукупність хімічних перетворень, спрямованих на збереження та самовідтворення біологічних систем.
Він включає надходження речовин в організм в процесі харчування і дихання, внутрішньоклітинний обмін речовин, або метаболізм, а також виділення кінцевих продуктів обміну.
Обмін речовин нерозривно пов'язані з процесами перетворення одних видів енергії на інші. Наприклад, у процесі фотосинтезу світлова енергія запасається як енергії хімічних зв'язків складних органічних молекул, а процесі дихання вона вивільняється і витрачається синтез нових молекул, механічну і осмотичну роботу, розсіюється як тепла тощо.
Перебіг хімічних реакцій у живих організмах забезпечується завдяки біологічним каталізаторам білкової природи. ферментам, або ензимам. Як і інші каталізатори, ферменти прискорюють протікання хімічних реакцій у клітині в десятки і сотні тисяч разів, а іноді й взагалі роблять їх можливими, але не змінюють при цьому ні природи, ні властивостей кінцевого продукту (продуктів) реакції та не змінюються самі. Ферменти можуть бути як простими, так і складними білками, до складу яких крім білкової частини входить і небілкова. кофактор (кофермент). Прикладами ферментів є амілаза слини, що розщеплює полісахариди при тривалому пережовуванні, та пепсин, що забезпечує перетравлення білків у шлунку.
p align="justify"> Ферменти відрізняються від каталізаторів небілкової природи високою специфічністю дії, значним збільшенням за їх допомогою швидкості реакції, а також можливістю регуляції дії за рахунок зміни умов протікання реакції або взаємодії з ними різних речовин. До того ж і умови, в яких протікає ферментний каталіз, суттєво відрізняються від тих, за яких йде неферментний: оптимальною для функціонування ферментів в організмі людини є температура $37°С$, тиск має бути близьким до атмосферного, а $рН$ середовища може суттєво вагатися. Так, для амілази необхідне лужне середовище, а для пепсину - кисле.
Механізм дії ферментів полягає у зниженні енергії активації речовин (субстратів), що вступають у реакцію, за рахунок утворення проміжних фермент-субстратних комплексів.

Енергетичний та пластичний обмін, їх взаємозв'язок
Метаболізм складається з двох процесів, що одночасно протікають у клітині: пластичного та енергетичного обмінів.
Пластичний обмін (анаболізм, асиміляція)являє собою сукупність реакцій синтезу, що йдуть із витратою енергії АТФ. У процесі пластичного обміну синтезуються органічні речовини, необхідні клітині. Прикладом реакцій пластичного обміну є фотосинтез, біосинтез білка та реплікація (самоподвоєння) ДНК.
Енергетичний обмін (катаболізм, дисиміляція)це сукупність реакцій розщеплення складних речовин до більш простих. Внаслідок енергетичного обміну виділяється енергія, що запасається у вигляді АТФ. Найбільш важливими процесами енергетичного обміну є дихання та бродіння.
Пластичний та енергетичний обміни нерозривно пов'язані, оскільки в процесі пластичного обміну синтезуються органічні речовини і для цього необхідна енергія АТФ, а в процесі енергетичного обміну органічні речовини розщеплюються та вивільняється енергія, яка потім буде витрачена на процеси синтезу.
Енергію організми отримують у процесі харчування, а вивільняють її та переводять у доступну форму в основному в процесі дихання. За способом харчування всі організми поділяються на автотрофи та гетеротрофи. Автотрофиздатні самостійно синтезувати органічні речовини з неорганічних, а гетеротрофивикористовують виключно готові органічні речовини.
Стадії енергетичного обміну
Незважаючи на всю складність реакцій енергетичного обміну, його умовно поділяють на три етапи: підготовчий, анаеробний (безкисневий) та аеробний (кисневий).
на підготовчому етапімолекули полісахаридів, ліпідів, білків, нуклеїнових кислот розпадаються більш прості, наприклад, глюкозу, гліцерин і жирні кислоти, амінокислоти, нуклеотиди та інших.
Анаеробний етапенергетичного обміну супроводжується подальшим розщепленням мономерів органічних сполук до ще більш простих проміжних продуктів, наприклад піровиноградної кислоти, або пірувату. Він не вимагає присутності кисню, і для багатьох організмів, що мешкають в мулі боліт або кишечнику людини, є єдиним способом отримання енергії. Анаеробний етап енергетичного обміну протікає у цитоплазмі.
Безкисневому розщепленню можуть піддаватися різні речовини, проте часто субстратом реакцій виявляється глюкоза. Процес її безкисневого розщеплення називається гліколізом. При гліколізі молекула глюкози втрачає чотири атоми водню, тобто окислюється, при цьому утворюються дві молекули піровиноградної кислоти, дві молекули АТФ і дві молекули відновленого переносника водню.
$С_6Н_(12)О_6 + 2Н_3РО_4 + 2АДФ + 2НАД → 2С_3Н_4О_3 + 2АТФ + 2НАДН + Н^(+) + 2Н_2О$.
Утворення АТФ з АДФ відбувається внаслідок прямого перенесення фосфат-аніону з попередньо фосфорильованого цукру та називається субстратним фосфорилуванням.
Аеробний етапенергетичного обміну може відбуватися тільки в присутності кисню, при цьому проміжні сполуки, що утворилися в процесі безкисневого розщеплення, окислюються до кінцевих продуктів (вуглекислого газу та води) і виділяється більша частина енергії, запасеної в хімічних зв'язках органічних сполук. Вона перетворюється на енергію макроергічних зв'язків 36 молекул АТФ. Цей етап також називається тканинним диханням. У разі відсутності кисню проміжні сполуки перетворюються на інші органічні речовини, і цей процес називається бродінням.
Дихання
Механізм клітинного дихання схематично зображено на рис.

Аеробне дихання відбувається в мітохондріях, при цьому піровиноградна кислота спочатку втрачає один атом вуглецю, що супроводжується синтезом одного відновлювального еквівалента $НАДН + Н^(+)$ і молекули ацетилкоферменту А (ацетил-КоА):
$С_3Н_4О_3 + НАД + Н~КоА → СН_3СО~КоА + НАДН + Н^(+) + СО_2$.
Ацетил-КоА в матриксі мітохондрій залучається до ланцюга хімічних реакцій, сукупність яких називається циклом Кребса (циклом трикарбонових кислот, циклом лимонної кислоти). Під час цих перетворень утворюється дві молекули АТФ, ацетил-КоА повністю окислюється до вуглекислого газу, яке іони водню і електрони приєднуються до переносників водню $НАДН + Н^(+)$ і $ФАДН_2$. Переносники транспортують протони водню та електрони до внутрішніх мембран мітохондрій, що утворюють кристи. За допомогою білків-переносників протони водню нагнітаються в міжмембранний простір, а електрони передаються по так званому дихальному ланцюгу ферментів, розташованому на внутрішній мембрані мітохондрій, і скидаються на атоми кисню:
$O_2+2e^(-)→O_2^-$.
Слід зазначити, що деякі білки дихального ланцюга містять залізо та сірку.
З міжмембранного простору протони водню транспортуються назад в матрикс мітохондрій за допомогою спеціальних ферментів - АТФ-синтаз, а енергія, що виділяється при цьому, витрачається на синтез 34 молекул АТФ з кожної молекули глюкози. Цей процес називається окисним фосфорилюванням. У матриксі мітохондрій протони водню реагують з радикалами кисню з утворенням води:
$4H^(+)+O_2^-→2H_2O$.
Сукупність реакцій кисневого дихання може бути виражена так:
$2С_3Н_4О_3 + 6О_2 + 36Н_3РО_4 + 36АДФ → 6СО_2 + 38Н_2О + 36АТФ.$
Сумарне рівняння дихання виглядає таким чином:
$С_6Н_(12)О_6 + 6О_2 + 38Н_3РО_4 + 38АДФ → 6СО_2 + 40Н_2О + 38АТФ.$
Бродіння
За відсутності кисню або за його нестачі відбувається бродіння. Бродіння є еволюційно більш раннім способом отримання енергії, ніж дихання, проте воно енергетично менш вигідно, оскільки в результаті бродіння утворюються органічні речовини, все ще багаті на енергію. Розрізняють кілька основних видів бродіння: молочнокисле, спиртове, оцтовокисле та ін. Так, в скелетних м'язах без кисню в ході бродіння піровиноградна кислота відновлюється до молочної кислоти, при цьому відновні еквіваленти, що утворилися раніше, витрачаються, і залишаються всього дві молекули АТФ:
$2С_3Н_4О_3 + 2НАДН + Н^(+) → 2С_3Н_6О_3 + 2НАД$.
При бродінні за допомогою дріжджових грибів піровиноградна кислота в присутності кисню перетворюється на етиловий спирт та оксид вуглецю (IV):
$С_3Н_4О_3 + НАДН + Н^(+) → С_2Н_5ОН + СО_2 + НАД^(+)$.
При бродінні за допомогою мікроорганізмів з піровиноградної кислоти можуть утворитися також оцтова, олійна, мурашина кислоти та ін.
АТФ, отримана внаслідок енергетичного обміну, витрачається у клітині різні види роботи: хімічну, осмотичну, електричну, механічну і регуляторну. Хімічна робота полягає у біосинтезі білків, ліпідів, вуглеводів, нуклеїнових кислот та інших життєво важливих сполук. До осмотичної роботи відносять процеси поглинання клітиною та виведення з неї речовин, які у позаклітинному просторі знаходяться у концентраціях більших, ніж у самій клітині. Електрична робота тісно взаємопов'язана з осмотичною, оскільки саме в результаті переміщення заряджених частинок через мембрани формується заряд мембрани та набуваються властивості збудливості та провідності. Механічна робота пов'язана з рухом речовин та структур усередині клітини, а також клітини загалом. До регуляторної роботи відносять всі процеси, створені задля координацію процесів у клітині.
Фотосинтез, його значення, космічна роль
Фотосинтезомназивають процес перетворення енергії світла на енергію хімічних зв'язків органічних сполук за участю хлорофілу.
В результаті фотосинтезу утворюється близько 150 млрд. тонн органічної речовини і приблизно 200 млрд. тонн кисню щорічно. Цей процес забезпечує кругообіг вуглецю в біосфері, не даючи накопичуватися вуглекислому газу і перешкоджаючи тим самим виникненню парникового ефекту та перегріву Землі. Органічні речовини, що утворюються в результаті фотосинтезу, не витрачаються іншими організмами повністю, значна їх частина протягом мільйонів років утворила поклади корисних копалин (кам'яного та бурого вугілля, нафти). Останнім часом як паливо почали використовувати також ріпакову олію («біодизель») та спирт, отриманий із рослинних залишків. З кисню під впливом електричних розрядів утворюється озон, який формує озоновий екран, який захищає живе Землі від згубної дії ультрафіолетових променів.
Наш співвітчизник, видатний фізіолог рослин К. А. Тімірязєв (1843-1920) назвав роль фотосинтезу «космічної», оскільки він пов'язує Землю із Сонцем (космосом), забезпечуючи приплив енергії на планету.
Фази фотосинтезу. Світлові та темнові реакції фотосинтезу, їх взаємозв'язок
В 1905 англійський фізіолог рослин Ф. Блекмен виявив, що швидкість фотосинтезу не може збільшуватися безмежно, якийсь фактор обмежує її. На підставі цього він висунув припущення про наявність двох фаз фотосинтезу: світловийі темновий. При низькій інтенсивності освітлення швидкість світлових реакцій зростає пропорційно до наростання сили світла, і, крім того, дані реакції не залежать від температури, оскільки для їх протікання не потрібні ферменти. Світлові реакції протікають на мембранах тилакоїд.
Швидкість темнових реакцій, навпаки, зростає з підвищенням температури, проте після досягнення температурного порога $30°С цей зростання припиняється, що свідчить про ферментативний характер зазначених перетворень, що відбуваються в стромі. Слід зазначити, що світло також впливає на темнові реакції певний вплив, незважаючи на те, що вони називаються темновими.

Світлова фаза фотосинтезу протікає на мембранах тилакоїдів, що несуть кілька типів білкових комплексів, основними з яких є фотосистеми I та II, а також АТФсинтазу. До складу фотосистем входять пігментні комплекси, в яких, крім хлорофілу, є і каротиноїди. Каротиноїди вловлюють світло у тих галузях спектра, у яких цього не робить хлорофіл, а також захищають хлорофіл від руйнування світлом високої інтенсивності.
Крім пігментних комплексів, фотосистеми включають і низку білків-акцепторів електронів, які послідовно передають один одному електрони від молекул хлорофілу. Послідовність цих білків називається електротранспортним ланцюгом хлоропластів.
З фотосистемою II також асоційовано спеціальний комплекс білків, що забезпечує виділення кисню у процесі фотосинтезу. Цей кисневиділяючий комплекс містить іони марганцю та хлору.
У світловий фазікванти світла, або фотони, які потрапляють на молекули хлорофілу, розташовані на мембранах тилакоїдів, переводять їх у збуджений стан, що характеризується вищою енергією електронів. При цьому збуджені електрони від хлорофілу фотосистеми I передаються через ланцюг посередників на переносник водню НАДФ, який при цьому приєднує протони водню, що завжди є у водному розчині:
$ НАДФ + 2e^(-) + 2H^(+) → НАДФН + Н^(+)$.
Відновлений $НАДФН + Н^(+)$ буде згодом використаний у темновій стадії. Електрони від хлорофілу фотосистеми II також передаються по електронтранспортному ланцюзі, проте вони заповнюють «електронні дірки» хлорофілу фотосистеми I. Нестача електронів у хлорофілі фотосистеми II заповнюється за рахунок забирання у молекул води, яке відбувається за участю згадуваного вище кисневиділяючого комплексу. В результаті розкладання молекул води, яке називається фотолізом, утворюються протони водню та виділяється молекулярний кисень, що є побічним продуктом фотосинтезу:
$H_2O → 2H^(+) + 2e^(-) + (1)/(2)O_2$.
Генетична інформація у клітині. Гени, генетичний код та його властивості. Матричний характер реакцій біосинтезу. Біосинтез білка та нуклеїнових кислот
Генетична інформація у клітці
Відтворення собі подібних є одним із фундаментальних властивостей живого. Завдяки цьому явищу існує подібність як між організмами, а й між окремими клітинами, і навіть їх органоїдами (мітохондріями і пластидами). Матеріальною основою цієї подібності є передача зашифрованої в послідовності нуклеотидів ДНК генетичної інформації, що здійснюється завдяки процесам реплікації (самоподвоєння) ДНК. Реалізуються всі ознаки та властивості клітин та організмів завдяки білкам, структуру яких насамперед і визначають послідовності нуклеотидів ДНК. Тому першорядне значення у процесах метаболізму грає саме біосинтез нуклеїнових кислот та білка. Структурною одиницею спадкової є ген.
Гени, генетичний код та його властивості
Спадкова інформація в клітці не є монолітною, вона розбита на окремі слова - гени.
Ген- Це елементарна одиниця генетичної інформації.
Роботи за програмою «Геном людини», які проводилися одночасно в кількох країнах і були завершені на початку нинішнього століття, дали нам розуміння того, що у людини всього близько 25-30 тис. генів, але інформація з більшої частини нашої ДНК не зчитується ніколи, оскільки в ній міститься безліч безглуздих ділянок, повторів і генів, що кодують ознаки, що втратили значення для людини (хвіст, оволосіння тіла та ін). Крім того, було розшифровано низку генів, які відповідають за розвиток спадкових захворювань, а також генів-мішеней лікарських препаратів. Однак практичне застосування результатів, отриманих в ході реалізації цієї програми, відкладається доти, поки не будуть розшифровані геноми більшої кількості людей і стане зрозуміло, чим же вони різняться.
Гени, що кодують первинну структуру білка, рибосомальної чи транспортної РНК називаються структурними, а гени, які забезпечують активацію або придушення зчитування інформації зі структурних генів, регуляторними. Проте, навіть структурні гени містять регуляторні ділянки.
Спадкова інформація організмів зашифрована в ДНК у вигляді певних поєднань нуклеотидів та їх послідовності. генетичного коду. Його властивостями є: триплетність, специфічність, універсальність, надмірність та неперекриваність. Крім того, в генетичному коді відсутні розділові знаки.
Кожна амінокислота закодована в ДНК трьома нуклеотидами. триплетом,наприклад, метіонін закодований триплетом ТАЦ, тобто код триплетен. З іншого боку, кожен триплет кодує лише одну амінокислоту, у чому його специфічність чи однозначність. Генетичний код універсальний для всіх живих організмів, тобто спадкова інформація про білки людини може зчитуватися бактеріями і навпаки. Це свідчить про єдність походження органічного світу. Однак 64 комбінаціям нуклеотидів по три відповідає лише 20 амінокислот, внаслідок чого одну амінокислоту може кодувати 2-6 триплетів, тобто генетичний код надмірний, або вироджений. Три триплети не мають відповідних амінокислот, їх називають стоп-кодонамиоскільки вони позначають закінчення синтезу поліпептидного ланцюга.
Послідовність основ у триплетах ДНК та кодовані ними амінокислоти

*Стоп-кодон, що означає кінець синтезу поліпептидного ланцюга.
Скорочення назв амінокислот:
Ала - аланін
Арг - аргінін
Асн - аспарагін
АСП - аспарагінова кислота
Вал - валін
Гіс - гістидин
Глі - гліцин
Глн - глутамін
Глу - глутамінова кислота
Іле - ізолейцин
Лій - лейцин
Ліз - лізин
Мет - метіонін
Про - пролін
Сір - серін
Тир - тирозин
Тре - треонін
Три - триптофан
Фен - фенілаланін
Цис - цистеїн
Якщо почати зчитування генетичної інформації не з першого нуклеотиду в триплеті, а з другого, то станеться не лише зсув рамки зчитування — синтезований таким чином білок буде зовсім іншим не лише за послідовністю нуклеотидів, а й за структурою та властивостями. Між триплетами відсутні будь-які розділові знаки, тому немає жодних перешкод для зсуву рамки зчитування, що відкриває простір для виникнення і збереження мутацій.
Матричний характер реакцій біосинтезу
Клітини бактерій здатні подвоюватися кожні 20-30 хвилин, а клітини еукаріотів — щодня і навіть частіше, що потребує високої швидкості та точності реплікації ДНК. Крім того, кожна клітина містить сотні та тисячі копій багатьох білків, особливо ферментів, отже, для їх відтворення неприйнятний «штучний» спосіб їх виробництва. Більш прогресивним способом є штампування, яке дозволяє отримати численні точні копії продукту і, до того ж, знизити його собівартість. Для штампування необхідна матриця, з якою здійснюється відбиток.
У клітинах принцип матричного синтезу полягає в тому, що нові молекули білків і нуклеїнових кислот синтезуються відповідно до програми, закладеної в структурі молекул, що раніше існували, тих же нуклеїнових кислот (ДНК або РНК).
Біосинтез білка та нуклеїнових кислот
Реплікація ДНК.ДНК є дволанцюжковим біополімером, мономерами якого є нуклеотиди. Якби біосинтез ДНК відбувався за принципом ксерокопіювання, то неминуче виникали численні спотворення та похибки у спадковій інформації, які зрештою призвели б до загибелі нових організмів. Тому процес подвоєння ДНК відбувається іншим, напівконсервативним способом: молекула ДНК розплітається, і на кожному з ланцюгів синтезується новий ланцюг за принципом комплементарності. Процес самовідтворення молекули ДНК, що забезпечує точне копіювання спадкової інформації та передачу її з покоління до покоління, називається реплікацією(Від лат. реплікація- Повторення). Внаслідок реплікації утворюються дві абсолютно точні копії материнської молекули ДНК, кожна з яких несе по одній копії материнської.

Процес реплікації насправді вкрай складний, оскільки у ньому бере участь низку білків. Одні з них розкручують подвійну спіраль ДНК, інші розривають водневі зв'язки між нуклеотидами комплементарних ланцюгів, треті (наприклад, фермент ДНК-полімераза) підбирають за принципом комплементарності нові нуклеотиди і т.д. новоствореним дочірнім клітинам.
Помилки в процесі реплікації виникають вкрай рідко, проте якщо вони і відбуваються, то дуже швидко усуваються як ДНК-полімеразами, так і спеціальними ферментами репарації, оскільки будь-яка помилка в послідовності нуклеотидів може призвести до незворотної зміни структури та функцій білка і, зрештою, несприятливо позначитися на життєздатності нової клітини чи навіть особини.
Біосинтез білка.Як образно висловився видатний філософ ХІХ століття Ф. Енгельс: «Життя є форма існування білкових тіл». Структура і властивості білкових молекул визначаються їхньою первинною структурою, тобто послідовністю амінокислот, зашифрованою в ДНК. Від точності відтворення цієї інформації залежить як існування самого поліпептиду, а й функціонування клітини загалом, тому процес синтезу білка має значення. Він, мабуть, є найскладнішим процесом синтезу у клітині, оскільки тут бере участь до трьохсот різних ферментів та інших макромолекул. Крім того, він протікає з високою швидкістю, що потребує ще більшої точності.
У біосинтезі білка виділяють два основні етапи: транскрипцію та трансляцію.
Транскрипція(Від лат. транскрипціо- Переписування) - це біосинтез молекул іРНК на матриці ДНК.

Оскільки молекула ДНК містить два антипаралельних ланцюга, то зчитування інформації з обох ланцюгів призвело б до утворення абсолютно різних іРНК, тому їхній біосинтез можливий тільки на одному з ланцюгів, який називають кодуючим, або кодогенним, на відміну від другого, некодуючого, або некодогенного. Забезпечує процес переписування спеціальний фермент РНК-полімеразу, який підбирає нуклеотиди РНК за принципом комплементарності. Цей процес може протікати як в ядрі, так і в органоїдах, що мають власну ДНК, - мітохондріях та пластидах.
Синтезовані в процесі транскрипції молекули іРНК проходять складний процес підготовки до трансляції (мітохондріальні та пластидні іРНК можуть залишатися всередині органоїдів, де відбувається другий етап біосинтезу білка). У процесі дозрівання іРНК до неї приєднуються перші три нуклеотиди (АУГ) та хвіст з аденілових нуклеотидів, довжина якого визначає, скільки копій білка може синтезуватися на цій молекулі. Тільки потім зрілі іРНК залишають ядро через ядерні пори.
Паралельно у цитоплазмі відбувається процес активації амінокислот, у ході якого амінокислота приєднується до відповідної вільної тРНК. Цей процес каталізується спеціальним ферментом, нею витрачається АТФ.
Трансляція(Від лат. трансляціо– передача) – це біосинтез поліпептидного ланцюга на матриці іРНК, при якому відбувається переведення генетичної інформації в послідовність амінокислот поліпептидного ланцюга.

Другий етап синтезу білка найчастіше відбувається у цитоплазмі, наприклад на шорсткої ЕПС. Для його протікання необхідні наявність рибосом, активація тРНК, у ході якої вони приєднують відповідні амінокислоти, присутність іонів Mg2+, а також оптимальні умови середовища (температура, рН, тиск тощо).
Для початку трансляції ( ініціації) до готової до синтезу молекули іРНК приєднується мала субодиниця рибосоми, а потім за принципом комплементарності до першого кодону (АУГ) підбирається тРНК, що несе амінокислоту метіонін. Лише після цього приєднується велика субодиниця рибосоми. У межах зібраної рибосоми виявляються два кодони іРНК, перший із яких вже зайнятий. До сусіднього з ним кодону приєднується друга тРНК, що також несе амінокислоту, після чого між залишками амінокислот за допомогою ферментів утворюється пептидна зв'язок. Рибосома пересувається однією кодон иРНК; перша з тРНК, що звільнилася від амінокислоти, повертається в цитоплазму за наступною амінокислотою, а фрагмент майбутнього поліпептидного ланцюга як би повисає на тРНК, що залишилася. До нового кодону, що опинився в межах рибосоми, приєднується наступна тРНК, процес повторюється і крок за кроком поліпептидний ланцюг подовжується, тобто відбувається його елонгація.
Закінчення синтезу білка ( термінація) відбувається, як тільки в молекулі іРНК зустрінеться специфічна послідовність нуклеотидів, яка не кодує амінокислоту (стоп-кодон). Після цього рибосома, іРНК і поліпептидний ланцюг поділяються, а знову синтезований білок набуває відповідної структури і транспортується в ту частину клітини, де виконуватиме свої функції.
Трансляція є дуже енергоємним процесом, оскільки на приєднання однієї амінокислоти до тРНК витрачається енергія однієї молекули АТФ, ще кілька використовуються для просування рибосоми молекулою іРНК.
Для прискорення синтезу певних білкових молекул до молекули іРНК можуть послідовно приєднуватися кілька рибосом, які утворюють єдину структуру. полісом.
Клітина – генетична одиниця живого. Хромосоми, їх будова (форма та розміри) та функції. Число хромосом та їх видова сталість. Соматичні та статеві клітини. Життєвий цикл клітини: інтерфаза та мітоз. Мітоз - розподіл соматичних клітин. Мейоз. Фази мітозу та мейозу. Розвиток статевих клітин у рослин та тварин. Розподіл клітини - основа зростання, розвитку та розмноження організмів. Роль мейозу та мітозу
Клітина – генетична одиниця живого
Незважаючи на те, що нуклеїнові кислоти є носієм генетичної інформації, реалізація цієї інформації неможлива поза клітиною, що легко доводиться на прикладі вірусів. Дані організми, що містять часто лише ДНК або РНК, не можуть самостійно відтворюватися, для цього вони повинні використовувати спадковий апарат клітини. Навіть проникнути в клітину без допомоги самої клітини вони не можуть, крім використання механізмів мембранного транспорту або завдяки пошкодженню клітин. Більшість вірусів є нестабільними, вони гинуть вже після декількох годин перебування на відкритому повітрі. Отже, клітина є генетичною одиницею живого, що має мінімальний набір компонентів для збереження, зміни та реалізації спадкової інформації, а також її передачі нащадкам.
Більша частина генетичної інформації еукаріотичної клітини зосереджена у ядрі. Особливістю її організації є те, що, на відміну від ДНК прокаріотичної клітини, молекули ДНК еукаріотів не замкнуті і утворюють складні комплекси з білками - хромосоми.
Хромосоми, їх будова (форма та розміри) та функції
Хромосома(Від грец. хрому- колір, забарвлення та сома- Тіло) - це структура клітинного ядра, яка містить гени і несе певну спадкову інформацію про ознаки і властивості організму.
Іноді хромосомами називають кільцеві молекули ДНК прокаріотів. Хромосоми здатні до самоподвоєння, вони мають структурну і функціональну індивідуальність і зберігають її в ряді поколінь. Кожна клітина несе всю спадкову інформацію організму, але у ній працює лише невелика частина.
Основою хромосоми є дволанцюжкова молекула ДНК, упакована з білками. У еукаріотів з ДНК взаємодіють гістонові та негістонові білки, тоді як у прокаріотів гістонові білки відсутні.
Найкраще хромосоми видно під світловим мікроскопом у процесі поділу клітини, коли вони в результаті ущільнення набувають вигляду паличкоподібних тілець, розділених первинною перетяжкою. центромірою — на плечі. На хромосомі може бути також вторинна перетяжка, яка в деяких випадках відокремлює від основної частини хромосоми так званий супутник. Кінцеві ділянки хромосом називаються тіломірами. Теломери перешкоджають злипання кінців хромосом і забезпечують їх прикріплення до оболонки ядра в клітині, що не ділиться. На початку розподілу хромосоми подвоєні і складаються з двох дочірніх хромосом. хроматид, скріплених у центромірі.

За формою розрізняють рівноплечі, нерівноплечі та паличкоподібні хромосоми. Розміри хромосом суттєво варіюють, проте середня хромосома має розміри 5$×$1,4 мкм.
У деяких випадках хромосоми в результаті численних подвій ДНК містять сотні і тисячі хроматид: такі гігантські хромосоми називаються політенними. Вони зустрічаються у слинних залозах личинок дрозофіли, а також у травних залозах аскариди.
Число хромосом та їх видова сталість. Соматичні та статеві клітини
Відповідно до клітинної теорії клітина є одиницею будови, життєдіяльності та розвитку організму. Таким чином, такі найважливіші функції живого, як зростання, розмноження та розвиток організму забезпечуються на клітинному рівні. Клітини багатоклітинних організмів можна розділити на соматичні та статеві.
Соматичні клітини- це всі клітини тіла, що утворюються внаслідок мітотичного розподілу.
Вивчення хромосом дозволило встановити, що з соматичних клітин організму кожного біологічного виду характерне постійне число хромосом. Наприклад, у людини 46. Набір хромосом соматичних клітин називають диплоїдним(2n), або подвійним.

Статеві клітини, або гамети, - це спеціалізовані клітини, що служать для статевого розмноження.
У гаметах завжди міститься вдвічі менше хромосом, ніж у соматичних клітинах (у людини — 23), тому набір хромосом статевих клітин називається гаплоїдним(n), або одинарним. Його утворення пов'язане з мейотичним поділом клітини.
Кількість ДНК соматичних клітин позначається як 2c, а статевих – 1с. Генетична формула соматичних клітин записується як 2n2c, а статевих – 1n1с.
У ядрах деяких соматичних клітин кількість хромосом може відрізнятися від їхньої кількості в соматичних клітинах. Якщо ця відмінність більше на один, два, три і т. д. гаплоїдного набору, то такі клітини називають поліплоїдними(три-, тетра-, пентаплоїдними відповідно). У таких клітинах процеси метаболізму протікають зазвичай дуже інтенсивно.
Кількість хромосом сама по собі не є видоспецифічною ознакою, оскільки різні організми можуть мати однакову кількість хромосом, а споріднені різні. Наприклад, у малярійного плазмодія та кінської аскариди по дві хромосоми, а у людини та шимпанзе – 46 та 48 відповідно.
Хромосоми людини поділяються на дві групи: аутосоми та статеві хромосоми (гетерохромосоми). Автосому соматичних клітинах людини налічується 22 пари, вони однакові для чоловіків та жінок, а статевих хромосомтільки одна пара, але саме вона визначає стать особини. Існує два види статевих хромосом – X та Y. Клітини тіла жінки несуть по дві X-хромосоми, а чоловіків – X та Y.
Каріотип- це сукупність ознак хромосомного набору організму (число хромосом, їх форма та величина).
Умовний запис каріотипу включає загальну кількість хромосом, статеві хромосоми та можливі відхилення в наборі хромосом. Наприклад, каріотип нормального чоловіка записується як 46, XY, а каріотип нормальної жінки – 46, XX.
Життєвий цикл клітини: інтерфаза та мітоз
Клітини не виникають щоразу заново, вони утворюються лише внаслідок поділу материнських клітин. Після поділу дочірнім клітинам потрібен деякий час для формування органоїдів та придбання відповідної структури, яка б забезпечила виконання певної функції. Цей відрізок часу називається дозріванням.
Проміжок часу від появи клітини в результаті поділу до її поділу або загибелі називається життєвим циклом клітини.
У еукаріотичних клітин життєвий цикл поділяється на дві основні стадії: інтерфазу та мітоз.
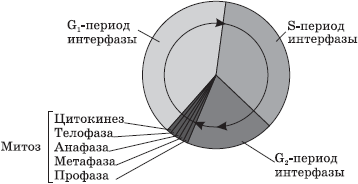
Інтерфаза- Це проміжок часу в життєвому циклі, в який клітина не ділиться та нормально функціонує. Інтерфаза ділиться на три періоди: G1-, S-і G2-періоди.
G 1 -період(пресинтетичний, постмітотичний) - це період росту та розвитку клітини, в який відбувається активний синтез РНК, білків та інших речовин, необхідних для повного життєзабезпечення клітини, що знову утворилася. До кінця цього періоду клітина може почати готуватися до подвоєння ДНК.
У S-період(Синтетичному) відбувається сам процес реплікації ДНК. Єдиною ділянкою хромосоми, яка не піддається реплікації, є центроміра, тому молекули ДНК, що утворилися, не розходяться повністю, а залишаються скріпленими в ній, і на початку поділу хромосома має X-подібний вигляд. Генетична формула клітини після подвоєння ДНК – 2n4c. Також у S-періоді відбувається подвоєння центріолей клітинного центру.
G 2 -період(постсинтетичний, премітотичний) характеризується інтенсивним синтезом РНК, білків та АТФ, необхідних для процесу поділу клітини, а також поділом центріолей, мітохондрій та пластид. До кінця інтерфази хроматин і ядерце залишаються добре помітними, цілісність ядерної оболонки не порушується, а органоїди не змінюються.
Частина клітин організму здатна виконувати свої функції протягом усього життя організму (нейрони нашого головного мозку, м'язові клітини серця), інші існують нетривалий час, після чого гинуть (клітини кишкового епітелію, клітини епідермісу шкіри). Отже, в організмі повинні постійно відбуватися процеси поділу клітин та утворення нових, які б заміщали відмерлі. Клітини, здатні до поділу, називають стволовими. В організмі людини вони знаходяться у червоному кістковому мозку, у глибоких шарах епідермісу шкіри та інших місцях. Використовуючи ці клітини, можна виростити новий орган, домогтися омолодження, і навіть клонувати організм. Перспективи використання стовбурових клітин цілком зрозумілі, проте морально-етичні аспекти цієї проблеми все ще обговорюються, оскільки в більшості випадків використовуються ембріональні стовбурові клітини, отримані з убитих при аборті зародків людини.
Тривалість інтерфази у клітинах рослин та тварин становить у середньому 10-20 годин, тоді як мітоз займає близько 1-2 годин.
У ході послідовних поділів у багатоклітинних організмах дочірні клітини стають дедалі різноманітнішими, оскільки у них відбувається зчитування інформації з дедалі більшої кількості генів.
Деякі клітини з часом перестають ділитися і гинуть, що може бути пов'язане із завершенням виконання певних функцій, як у разі клітин епідермісу шкіри та клітин крові або з пошкодженням цих клітин факторами довкілля, зокрема збудниками хвороб. Генетично запрограмована смерть клітини називається апоптозом, тоді як випадкова загибель некрозом.
Мітоз – розподіл соматичних клітин. Фази мітозу
Мітоз- Спосіб непрямого поділу соматичних клітин.
Під час мітозу клітина проходить ряд послідовних фаз, у результаті яких кожна дочірня клітина отримує такий самий набір хромосом, як і материнській клітині.
Мітоз ділиться на чотири основні фази: профазу, метафазу, анафазу та телофазу. Профаза— найбільш тривала стадія мітозу, у процесі якої відбувається конденсація хроматину, у результаті стають видні X-подібні хромосоми, які з двох хроматид (дочірніх хромосом). При цьому зникає ядерце, центріолі розходяться до полюсів клітини, і починає формуватися ахроматинове веретено (веретено поділу) з мікротрубочок. Наприкінці профази ядерна оболонка розпадається окремі бульбашки.

У метафазіхромосоми вишиковуються за екватором клітини своїми центромірами, до яких прикріплюються мікротрубочки повністю сформованого веретена поділу. На цій стадії розподілу хромосоми найбільш ущільнені та мають характерну форму, що дозволяє вивчити каріотип.
У анафазевідбувається швидка реплікація ДНК у центромірах, внаслідок якої хромосоми розщеплюються та хроматиди розходяться до полюсів клітини, що розтягуються мікротрубочками. Розподіл хроматид має бути абсолютно рівним, оскільки саме цей процес забезпечує підтримку сталості числа хромосом у клітинах організму.
На стадії телофазидочірні хромосоми збираються на полюсах, деспіралізуються, навколо них з бульбашок формуються ядерні оболонки, а в ядрах, що знову утворилися, виникають ядерця.
Після розподілу ядра відбувається розподіл цитоплазми. цитокінез,в ході якого і відбувається більш менш рівномірний розподіл всіх органоїдів материнської клітини.
Таким чином, в результаті мітозу з однієї материнської клітини утворюється дві дочірні, кожна з яких є генетичною копією материнської (2n2c).
У хворих, пошкоджених, старіючих клітинах та спеціалізованих тканинах організму може відбуватися дещо інший процес поділу – амітоз. Амітозомназивають пряме розподіл еукаріотичних клітин, у якому немає утворення генетично рівноцінних клітин, оскільки клітинні компоненти розподіляються нерівномірно. Він зустрічається у рослин в ендоспермі, а у тварин – у печінці, хрящах та рогівці ока.
Мейоз. Фази мейозу
Мейоз- це спосіб непрямого поділу первинних статевих клітин (2n2с), в результаті якого утворюються гаплоїдні клітини (1n1с), найчастіше статеві.
На відміну від мітозу, мейоз складається з двох послідовних поділів клітини, кожному з яких передує інтерфаза. Перший поділ мейозу (мейоз I) називається редукційнимоскільки при цьому кількість хромосом зменшується вдвічі, а друге поділ (мейоз II) екваційним, оскільки у процесі кількість хромосом зберігається.

Інтерфаза Iпротікає подібно до інтерфази мітозу. Мейоз Iділиться на чотири фази: профазу I, метафазу I, анафазу I та телофазу I. профазі Iвідбуваються два найважливіші процеси — кон'югація та кросинговер. Кон'югація- Це процес злиття гомологічних (парних) хромосом по всій довжині. Пари хромосом, що утворилися в процесі кон'югації, зберігаються до кінця метафази I.
Кросинговер- Взаємний обмін гомологічними ділянками гомологічних хромосом. В результаті кросинговера хромосоми, отримані організмом від обох батьків, набувають нових комбінацій генів, що зумовлює появу генетично різноманітного потомства. Наприкінці профази I, як і в профазі мітозу, зникає ядерце, центріолі розходяться до полюсів клітини, а ядерна оболонка розпадається.

У метафазі Iпари хромосом шикуються по екватору клітини, до їх центромірів прикріплюються мікротрубочки веретена поділу.
У анафазі Iдо полюсів розходяться цілі гомологічні хромосоми, які з двох хроматид.
У телофазі Iнавколо скупчень хромосом біля полюсів клітини утворюються ядерні оболонки, формуються ядерця.
Цитокінез Iзабезпечує розподіл цитоплазм дочірніх клітин.
Дочірні клітини (1n2c), що утворилися в результаті мейозу I, генетично різнорідні, оскільки їх хромосоми, що випадково розійшлися до полюсів клітини, містять неоднакові гени.
Порівняльна характеристика мітозу та мейозу
| Ознака | Мітоз | Мейоз | |
| Які клітини вступають у розподіл? | Соматичні (2n) | Первинні статеві клітини (2n) | |
| Число поділів | 1 | 2 | |
| Скільки та яких клітин утворюється у процесі поділу? | 2 соматичні (2n) | 4 статеві (n) | |
| Інтерфаза | Підготовка клітини до поділу, подвоєння ДНК | Дуже коротка, подвоєння ДНК не відбувається | |
| Фази | Мейоз I | Мейоз II | |
| Профаза | Конденсація хромосом, зникнення ядерця, розпад ядерної оболонки, можуть відбуватися кон'югація та кросинговер | Конденсація хромосом, зникнення ядерця, розпад ядерної оболонки | |
| Метафаза | За екватором розташовуються пари хромосом, формується веретено поділу | Хромосоми вишиковуються за екватором, формується веретено поділу | |
| Анафаза | До полюсів розходяться гомологічні хромосоми із двох хроматид. | До полюсів розходяться хроматиди | |
| Телофаза | Хромосоми деспіралізуються, формуються нові ядерні оболонки та ядерця | Хромосоми деспіралізуються, формуються нові ядерні оболонки та ядерця | |
Інтерфаза IIдуже коротка, оскільки у ній немає подвоєння ДНК, тобто відсутня S-період.
Мейоз IIтакож ділиться на чотири фази: профазу II, метафазу II, анафазу II та телофазу II. У профазі IIпротікають самі процеси, як у профазі I, крім кон'югації і кросинговера.
У метафазі IIХромосоми розташовуються вздовж екватора клітини.
У анафазе IIхромосоми розщеплюються в центромірах і до полюсів вже розтягуються хроматиди.
У телофазі IIнавколо скупчень дочірніх хромосом формуються ядерні оболонки та ядерця.
Після цитокінезу IIгенетична формула всіх чотирьох дочірніх клітин - 1n1c, проте всі вони мають різний набір генів, що є результатом кросинговеру та випадкового поєднання хромосом материнського та батьківського організмів у дочірніх клітинах.
Розвиток статевих клітин у рослин та тварин
Гаметогенез(Від грец. гамете- дружина, гаметес- Чоловік і генезис- Походження, виникнення) - це процес утворення зрілих статевих клітин.
Так як для статевого розмноження найчастіше необхідні дві особини - жіноча і чоловіча, які продукують різні статеві клітини - яйцеклітини та спермії, то і процеси утворення цих гамет мають бути різні.
Характер процесу істотно залежить і від того, чи відбувається він у рослинній або тваринній клітині, оскільки у рослин при утворенні гамет відбувається лише мітоз, а у тварин — і мітоз, і мейоз.
Розвиток статевих клітин рослин.У покритонасінних рослин утворення чоловічих і жіночих статевих клітин відбувається в різних частинах квітки - тичинках та маточках відповідно.
Перед утворенням чоловічих статевих клітин мікрогаметогенезом(Від грец. мікрос- маленький) - відбувається мікроспорогенез, тобто формування мікроспор у пильовиках тичинок. Цей процес пов'язаний з мейотичним розподілом материнської клітини, в результаті якого виникають чотири гаплоїдні мікроспори. Мікрогаметогенез пов'язаний з мітотичним розподілом мікроспори, що дає чоловічий гаметофіт із двох клітин - крупний вегетативною(сифоногенної) та дрібної генеративної. Після поділу чоловічий гаметофіт покривається щільними оболонками та утворює пилкове зерно. У деяких випадках ще в процесі дозрівання пилку, а іноді тільки після перенесення на рильці маточка генеративна клітина ділиться мітотично з утворенням двох нерухомих чоловічих статевих клітин. сперміїв. З вегетативної клітини після запилення формується пилкова трубка, за якою спермії проникають у зав'язь маточка для запліднення.

Розвиток жіночих статевих клітин у рослин називається мегагаметогенезом(Від грец. мегас- Великий). Він відбувається в зав'язі маточка, чому передує мегаспорогенез, в результаті якого з материнської клітини мегаспори, що лежить в нуцеллусі, шляхом мейотичного поділу формуються чотири мегаспори. Одна з мегаспор тричі ділиться мітотично, даючи жіночий гаметофіт — зародковий мішок із вісьма ядрами. При подальшому відокремленні цитоплазм дочірніх клітин одна з клітин, що утворилися, стає яйцеклітиною, з боків від якої лежать так звані синергіди, на протилежному кінці зародкового мішка формуються три антиподи, а в центрі в результаті злиття двох гаплоїдних ядер утворюється диплоїдна центральна клітина.

Розвиток статевих клітин у тварин.У тварин розрізняють два процеси утворення статевих клітин - сперматогенез та овогенез.
Сперматогенез(Від грец. сперма, сперматос- Насіння і генезис- Походження, виникнення) - це процес утворення зрілих чоловічих статевих клітин - сперматозоїдів. У людини він протікає в сім'яниках, або яєчках, і ділиться на чотири періоди: розмноження, зростання, дозрівання та формування.

У період розмноженняпервинні статеві клітини діляться мітотично, внаслідок чого утворюються диплоїдні сперматогонії. У період зростаннясперматогонії накопичують поживні речовини в цитоплазмі, збільшуються в розмірах і перетворюються на первинні сперматоцити, або сперматоцити 1-го порядку. Лише після цього вони вступають у мейоз ( період дозрівання), в результаті якого утворюється спочатку два вторинних сперматоцитів, або сперматоцита 2-го порядку, а потім - чотири гаплоїдні клітини з ще досить великою кількістю цитоплазми - сперматіди. У період формуваннявони втрачають майже всю цитоплазму і формують джгутик, перетворюючись на сперматозоїди.
Сперматозоїди, або живчики, - дуже дрібні рухливі чоловічі статеві клітини, що мають голівку, шийку та хвостик.

У голівцікрім ядра, знаходиться акросома- видозмінений комплекс Гольджі, що забезпечує розчинення оболонок яйцеклітини в процесі запліднення. У шийцізнаходяться центріолі клітинного центру, а основу хвостикаутворюють мікротрубочки, що безпосередньо забезпечують рух сперматозоїда. У ньому також розташовані мітохондрії, що забезпечують сперматозоїд енергією АТФ для руху.
Овогенез(Від грец. він- яйце і генезис- Походження, виникнення) - це процес утворення зрілих жіночих статевих клітин - яйцеклітин. У людини вона відбувається в яєчниках і складається з трьох періодів: розмноження, зростання та дозрівання. Періоди розмноження та зростання, аналогічні таким у сперматогенезі, відбуваються ще під час внутрішньоутробного розвитку. При цьому з первинних статевих клітин у результаті мітозу утворюються диплоїдні. оогонії, які перетворюються потім на диплоїдні первинні ооцити, або ооцити 1-го порядку. Мейоз і наступний цитокінез, що протікають період дозрівання, характеризуються нерівномірністю поділу цитоплазми материнської клітини, так що в результаті спочатку виходить один вторинний ооцит, або ооцит 2-го порядку, і перше полярне тільце, а потім із вторинного ооциту - яйцеклітина, що зберігає весь запас поживних речовин, і друге полярне тільце, тоді як перше полярне тільце ділиться на два. Полярні тільця забирають надлишок генетичного матеріалу.
У людини яйцеклітини виробляються із проміжком 28-29 діб. Цикл, пов'язаний із дозріванням та виходом яйцеклітин, називається менструальним.
Яйцеклітина— велика жіноча статева клітина, яка несе як гаплоїдний набір хромосом, а й значний запас поживних речовин подальшого розвитку зародка.

Яйцеклітина у ссавців покрита чотирма оболонками, що знижують ймовірність її пошкодження різними факторами. Діаметр яйцеклітини у людини досягає 150-200 мкм, тоді як у страуса він може становити кілька сантиметрів.
Розподіл клітини - основа зростання, розвитку та розмноження організмів. Роль мітозу та мейозу
Якщо в одноклітинних організмів розподіл клітини призводить до збільшення кількості особин, т. е. розмноження, то багатоклітинних цей процес може мати різне значення. Так, розподіл клітин зародка, починаючи з зиготи, є біологічною основою взаємопов'язаних процесів зростання та розвитку. Подібні зміни спостерігаються в людини в підлітковому віці, коли число клітин не тільки збільшується, але і відбувається якісна зміна організму. В основі розмноження багатоклітинних організмів також лежить поділ клітини, наприклад, при безстатевому розмноженні завдяки цьому процесу з частини організму відбувається відновлення цілісного, а при статевому — у процесі гаметогенезу утворюються статеві клітини, що дають згодом новий організм. Слід зазначити, основні способи поділу еукаріотичної клітини — мітоз і мейоз — мають різне значення у життєвих циклах організмів.
В результаті мітозу відбувається рівномірний розподіл спадкового матеріалу між дочірніми клітинами - точними копіями материнської. Без мітозу було б неможливим існування та зростання багатоклітинних організмів, що розвиваються з єдиної клітини – зиготи, оскільки всі клітини таких організмів мають містити однакову генетичну інформацію.
У процесі поділу дочірні клітини стають все більш різноманітними за будовою та виконуваними функціями, що пов'язано з активацією у них нових груп генів внаслідок міжклітинної взаємодії. Таким чином, мітоз необхідний розвитку організму.
Цей спосіб поділу клітин необхідний процесів безстатевого розмноження і регенерації (відновлення) пошкоджених тканин, і навіть органів.
Мейоз, у свою чергу, забезпечує сталість каріотипу при статевому розмноженні, оскільки зменшує вдвічі набір хромосом перед статевим розмноженням, який відновлюється в результаті запліднення. Крім того, мейоз призводить до появи нових комбінацій батьківських генів завдяки кросинговеру та випадковому поєднанню хромосом у дочірніх клітинах. Завдяки цьому потомство виходить генетично різноманітним, що дає матеріал для природного відбору та є матеріальною основою еволюції. Зміна числа, форми та розмірів хромосом, з одного боку, може призвести до появи різних відхилень у розвитку організму і навіть його загибелі, а з іншого - може призвести до появи особин, більш пристосованих до довкілля.
Таким чином, клітина є одиницею зростання, розвитку та розмноження організмів.
Третій етап еволюції – поява клітини.
Молекули білків та нуклеїнових кислот (ДНК та РНК) утворюють біологічну клітину, найменшу одиницю живого. Біологічні клітини є "будівельними цеглинами" всіх живих організмів і містять у собі всі матеріальні коди розвитку.
Довгий час вчені вважали влаштування клітини гранично простим. Радянський енциклопедичний словник трактує поняття клітини так: "Клітка - елементарна жива система, основа будови та життєдіяльності всіх тварин та рослин". Слід зазначити, що термін "елементарна" в жодному разі не означає "найпростіша".
Коли за допомогою електронного мікроскопа вдалося заглянути всередину, виявилося, що пристрій найпростішої клітини так само складно і незрозуміло, як сам Всесвіт. Сьогодні вже встановлено, що "Клітка - це особлива матерія Всесвіту, особлива матерія Космосу". Одна єдина клітина містить відомості, які можна укласти лише кілька десятків тисяч томів Великої радянської енциклопедії. Тобто. Клітина, крім усього іншого, - величезний "біорезервуар" інформації".
Автор сучасної теорії молекулярної еволюції Манфред Ейген пише: "Для того, щоб білкова молекула утворилася випадково, природі довелося б зробити приблизно 10130 проб і витратити на це таке число молекул, якого вистачило б на 1027 Всесвітів. Якщо ж білок будувався розумно, тобто так Що обґрунтованість кожного ходу могла бути перевірена якимось механізмом селекції, то на це знадобилося всього близько 2000 спроб.Ми приходимо до парадоксального висновку: програма побудови "первісної живої клітини" закодована десь на рівні елементарних частинок.
Та й як може бути інакше. Кожна клітина, володіючи ДНК, наділена свідомістю, усвідомлює себе та інші клітини, і перебуває в контакті з Всесвітом, будучи, по суті, її частиною. І хоча кількість і різноманітність клітин в організмі людини вражає (близько 70 трильйонів), всі вони самоподібні, як самоподібні всі процеси, що відбуваються в клітинах. За словами німецького вченого Роланда Глазера, конструкція біологічних клітин "дуже добре продумана". Ким добре продумана?
Відповідь проста: білки, нуклеїнові кислоти, живі клітини та всі біологічні системи є продуктом творчої діяльності інтелектуального Творця.
Що цікаво: на атомному рівні відмінностей між хімічним складом органічного та неорганічного світу немає. Іншими словами, на рівні атома клітина створена з тих самих елементів, що й нежива природа. Відмінності виявляються на молекулярному рівні. У живих тілах поряд з неорганічними речовинами та водою знаходяться ще білки, вуглеводи, жири, нуклеїнові кислоти, фермент АТФ-синтаза та інші низькомолекулярні органічні сполуки.
На сьогоднішній день клітину з метою вивчення буквально розібрали на атоми. Однак створити хоч одну живу клітину так і не вдається, бо створити клітину це означає створити частинку живого Всесвіту. Академік В.П. Казначеєв вважає, що "клітина - це космопланетарний організм… Клітини людини - це певні системи ефіроторсійних біоколайдерів. У цих біоколайдерах відбуваються невідомі нам процеси, йде матеріалізація космічних форм потоків, їх космоперетворення і за рахунок цього частки матералізуються".
Вода.
Майже 80 % маси клітини становить вода. За твердженням доктора біологічних наук С. Зеніна вода завдяки своїй кластерній структурі є інформаційною матрицею для управління біохімічними процесами. Крім того, саме вода є тією первинною "мішенню", з якою взаємодіють коливання звукової частоти. Впорядкованість клітинної води настільки висока (близька до впорядкованості кристала), що її називають рідким кристалом.
Білки.
Величезну роль біологічному житті грають білки. У клітині містяться кілька тисяч білків, властивих лише даному виду клітини (виняток становлять стовбурові клітини). Здатність синтезувати саме свої білки передається у спадок від клітини до клітини та зберігається протягом усього життя. У процесі життєдіяльності клітини білки поступово змінюють свою структуру, їхня функція порушується. Ці білки, що відпрацювали, видаляються з клітини і замінюються новими, завдяки чому життєдіяльність клітини зберігається.
Зазначимо, насамперед, будівельну функцію білків, бо вони є тим будівельним матеріалом, з якого складаються мембрани клітин і клітинних органоїдів, стінки кровоносних судин, сухожилля, хрящі тощо.
Надзвичайно цікава сигнальна функція білків. Виявляється, білки здатні служити сигнальними речовинами, передаючи сигнали між тканинами, клітинами чи організмами. Сигнальну функцію виконують білки-гормони. Клітини можуть взаємодіяти одна з одною на відстані за допомогою сигнальних білків, що передаються через міжклітинну речовину.
Білкам властива також рухова функція. Усі види руху, яких здатні клітини, наприклад, скорочення м'язів, виконують особливі скорочувальні білки. Білки виконують також транспортну функцію. Вони здатні приєднувати різні речовини та переносити їх з одного місця клітини до іншого. Наприклад, білок крові гемоглобін приєднує кисень і розносить його до всіх тканин та органів тіла. Крім того, білкам притаманна і захисна функція. При введенні чужорідних білків або клітин в організм у ньому відбувається вироблення особливих білків, які пов'язують та знешкоджують чужорідні клітини та речовини. Ну і нарешті, енергетична функція білків у тому, що з повному розщепленні 1г білка звільняється енергія у кількості 17, 6 кДж.
Будова клітини.
Клітина складається з трьох нерозривно пов'язаних між собою частин: оболонки, цитоплазми та ядра, причому будова та функція ядра у різні періоди життя клітини різні. Бо життя клітини включає два періоди: поділ, в результаті якого утворюються дві дочірні клітини, і період між поділами, який називається інтерфазою.
Оболонка клітини здійснює безпосередню взаємодію із зовнішнім середовищем та взаємодію із сусідніми клітинами. Вона складається із зовнішнього шару та розташованої під ним плазматичної мембрани. Поверхневий шар тварин клітин називається глікоколісом. Він здійснює зв'язок клітин із зовнішнім середовищем і з усіма оточуючими його речовинами. Товщина його не більше 1мкм.
Будова клітини
Клітинна мембрана – дуже важлива частина клітини. Вона утримує разом усі клітинні компоненти та розмежовує зовнішнє та внутрішнє середовище.
Між клітинами та зовнішнім середовищем постійно відбувається обмін речовин. З довкілля в клітину надходить вода, різноманітні солі у формі окремих іонів, неорганічні та органічні молекули. У зовнішнє середовище через мембрану з клітини виводяться продукти обміну, а також речовини, синтезовані у клітині: білки, вуглеводи, гормони, що виробляються у клітинах різних залоз. Транспорт речовин - одне з основних функцій плазматичної мембрани.
Цитоплазма- внутрішнє напіврідке середовище, в якому протікають основні процеси обміну речовин. Останні дослідження показали, що цитоплазма - це не є розчин, компоненти якого взаємодіють один з одним при випадкових зіткненнях. Її можна порівняти з желе, яке починає "тремтіти" у відповідь на зовнішній вплив. Саме так цитоплазма сприймає та передає інформацію.
У цитоплазмі розташовуються ядро та різні органоїди, що об'єднуються нею в одне ціле, що забезпечує їхню взаємодію та діяльність клітини як єдиної цілісної системи. Ядро розташовується у центральній частині цитоплазми. Вся внутрішня зона цитоплазми заповнена ендоплазматичною мережею, яка є клітинним органоїдом: система канальців, бульбашок і "цистерн", відмежованих мембранами. Ендоплазматична мережа бере участь в обмінних процесах, забезпечуючи транспорт речовин з навколишнього середовища до цитоплазми та між окремими внутрішньоклітинними структурами, але основна її функція – участь у синтезі білка, який здійснюється у рибосомах. - мікроскопічних тільцях округлої форми діаметром 15-20 нм. Синтезовані білки спочатку накопичуються в каналах та порожнинах ендоплазматичної мережі, а потім транспортуються до органоїдів та ділянок клітини, де вони споживаються.
Крім білків у цитоплазмі містяться також мітохондрії, дрібні тільця розміром 0,2-7мкм., які називають "силовими станціями" клітин. У мітохондріях протікають окислювально-відновні реакції, що забезпечують клітини енергією. Число мітохондрій в одній клітці від одиниць до кількох тисяч.
Ядро- життєва частина клітини, керує синтезом білків і крізь них усіма фізіологічними процесами у клітині. У ядрі клітини, що не розділяється, розрізняють ядерну оболонку, ядерний сік, ядерце і хромосоми. Через ядерну оболонку здійснюється безперервний обмін речовин між ядром та цитоплазмою. Під ядерною оболонкою - ядерний сік (напіврідка речовина), в якому знаходяться ядерце і хромосоми. Ядро - щільне округле тільце, розміри якого можуть змінюватися в широких межах, від 1 до 10 мкм і більше. Воно складається в основному з рибонуклеопротеїдів; бере участь у освіті рибосом. Зазвичай у клітині 1-3 ядерця, іноді до кількох сотень. До складу ядерця входять РНК та білок.
З появою клітки на Землі виникло Життя!
Далі буде...



